 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Digital biology and chemistry†
Daan
Witters
,
Bing
Sun
,
Stefano
Begolo
,
Jesus
Rodriguez-Manzano
,
Whitney
Robles
and
Rustem F.
Ismagilov
*
Division of Chemistry and Chemical Engineering, California Institute of Technology, 1200 E. California Blvd., Pasadena, CA 91125, USA. E-mail: rustem.admin@caltech.edu
First published on 2nd June 2014
Abstract
This account examines developments in “digital” biology and chemistry within the context of microfluidics, from a personal perspective. Using microfluidics as a frame of reference, we identify two areas of research within digital biology and chemistry that are of special interest: (i) the study of systems that switch between discrete states in response to changes in chemical concentration of signals, and (ii) the study of single biological entities such as molecules or cells. In particular, microfluidics accelerates analysis of switching systems (i.e., those that exhibit a sharp change in output over a narrow range of input) by enabling monitoring of multiple reactions in parallel over a range of concentrations of signals. Conversely, such switching systems can be used to create new kinds of microfluidic detection systems that provide “analog-to-digital” signal conversion and logic. Microfluidic compartmentalization technologies for studying and isolating single entities can be used to reconstruct and understand cellular processes, study interactions between single biological entities, and examine the intrinsic heterogeneity of populations of molecules, cells, or organisms. Furthermore, compartmentalization of single cells or molecules in “digital” microfluidic experiments can induce switching in a range of reaction systems to enable sensitive detection of cells or biomolecules, such as with digital ELISA or digital PCR. This “digitizing” offers advantages in terms of robustness, assay design, and simplicity because quantitative information can be obtained with qualitative measurements. While digital formats have been shown to improve the robustness of existing chemistries, we anticipate that in the future they will enable new chemistries to be used for quantitative measurements, and that digital biology and chemistry will continue to provide further opportunities for measuring biomolecules, understanding natural systems more deeply, and advancing molecular and cellular analysis. Microfluidics will impact digital biology and chemistry and will also benefit from them if it becomes massively distributed.
Introduction
Various modes of inquiry in biology and chemistry can be characterized as “digital.” Within the context of microfluidics, two broad areas of research are especially relevant and have shaped how the authors view the emerging disciplines of digital biology and chemistry: (i) the study of switching in natural systems (Fig. 1A) and (ii) the study of single biological entities (Fig. 1B). Combining the two with lab-on-a-chip technologies provides new opportunities for precisely measuring quantities of biomolecules, characterizing the mechanisms of natural systems, and advancing molecular and cellular analysis.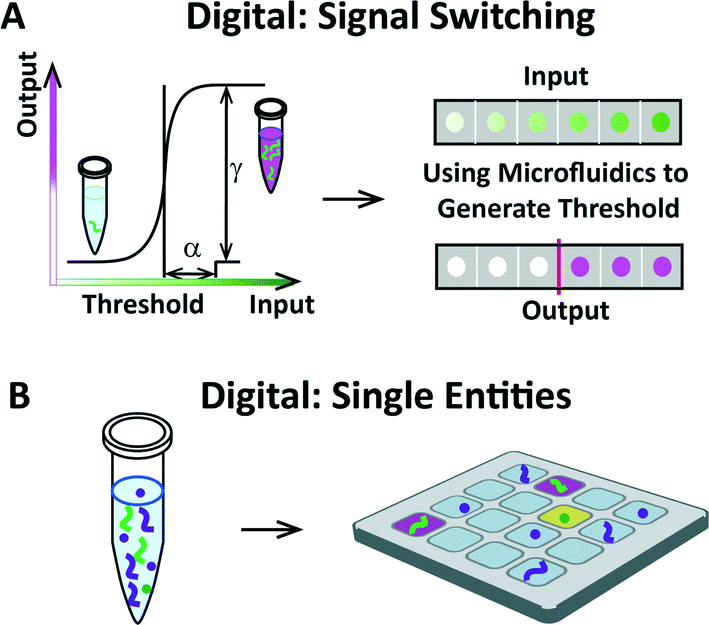 | ||
| Fig. 1 Schematic overview of switching systems and single entities, two areas of research in “digital” biology and chemistry that can be enhanced by using microfluidics. (A) Left: a graph illustrating signal switching, defined by a sharp, often sigmoidal change in output over a narrow range of input. In this example, switching occurs when the concentration of signals exceeds a certain threshold by a value of concentration equal to α, resulting in a drastic change in output given by the value of γ. Sharpness of switching can be defined via α and γ.2 Right: schematic showing a gradient range of input translated into digital output. When studied using microfluidics, switching systems can be precisely characterized. (B) Left: schematic of a tube containing a variety of single entities (such as molecules or cells). Right: schematic showing that single biological entities can be compartmentalized on a microfluidic device to enable simplified readout and analysis of compartment contents (for example, to enable studies of heterogeneity or signaling). | ||
The following personal account of digital biology and chemistry is not intended to be a comprehensive review of the literature; given the citation limit, we could not cover the full scope of exciting work in these fields, and we regret that we could not cover excellent work by many friends and colleagues who have made foundational contributions to these fields. For instance, we do not review all technologies that have been characterized as “digital microfluidics”, such as electrowetting, as they have been extensively treated in other reviews.1 Rather, we focus on digital biology and chemistry in the context of the two areas of research mentioned above and describe these digital approaches primarily in terms of how we have experienced and approached them through our own research.
“Digital” as switching
In chemistry and biology, “digital” can refer to the tendency of many natural systems to switch between discrete states (e.g., off or on, clotted or not clotted, activated or not, differentiated or not) in response to specific signals such as proteins, carbohydrates, nucleic acids, autoinducers, or redox potential gradients. Such switching may apply to entire reactions and their networks as well as to individual cells. This is distinct from but analogous to the use of the word “digital” in computing, where it refers to the binary code of 0s and 1s on which computers operate.In biochemical systems, switching is defined by a sharp, often sigmoidal change in output over a narrow range of input (Fig. 1A). This change may result from the accumulation of a specific signal above a threshold concentration or the accumulation of a specific event over time (e.g., DNA mutations) that will increase the fitness of one population, therefore leading to switching under selective pressure in a given environment.
Such switching in nature is widespread and well known, and it plays many roles in natural systems (Fig. 2). Among the simple switching systems we find examples such as inducible operons (e.g., the lac operon is normally “off” but turns “on” in the presence of an inducer, lactose)8 and co-repressor proteins (e.g., the transcriptional repressor MBD2, which has been reported to be associated to the transcriptional repressor complex composed by HDAC and MeCP1).9 In these cases, when the signal exceeds a certain threshold concentration, one single reaction occurs and switches the state of the system. In more complicated systems, hundreds of genes and proteins must be coordinated and regulated: one or more signals first stimulate a certain response, which further induces more downstream reactions and eventually determines the fate of the system as a whole. Examples include (i) genetic regulatory networks, where a subset of transcription factors connect signaling pathways with genes to produce the appropriate gene activities,6 (ii) cell cycle control, where an ordered set of events culminates in cell growth and division into two daughter cells,4 (iii) cancer, where the proteins involved in regulating cell division events no longer appropriately drive progression from one cell cycle stage to the next,10 and (iv) the adaptive immune system, which is activated by the innate response to protect the host against pathogens.11 Switching between two states plays an important role in natural systems both as an indicator of the physical location of certain biological events, such as in the case of bacterial quorum sensing,3 and as the basic unit of decision-making in an organism's development.
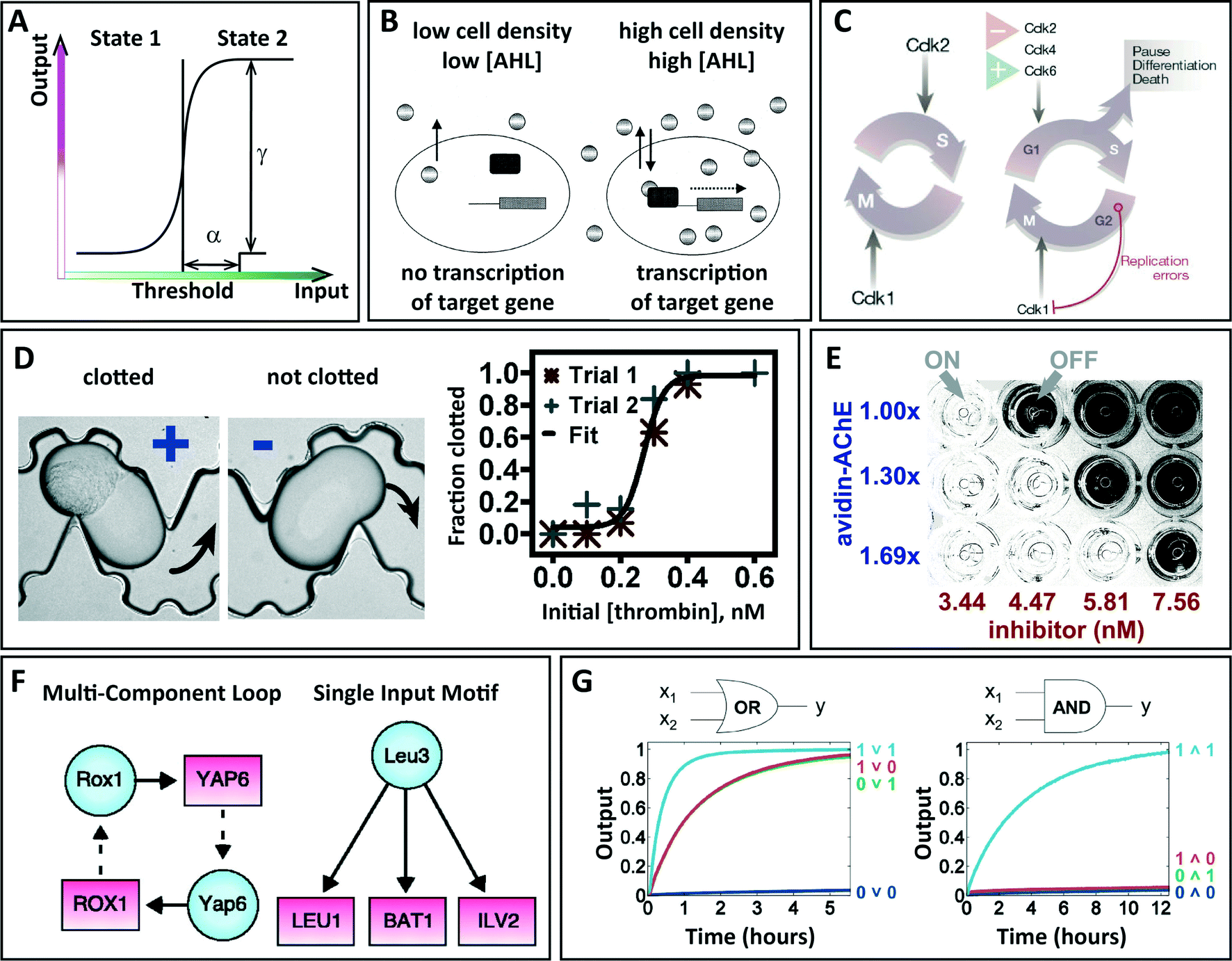 | ||
| Fig. 2 Examples of switching systems in nature, uses of microfluidics to study switching, and future applications. (A) Graphical definition of switching. (B–C) Natural examples of switching. (B) An illustration of bacterial quorum sensing in gram-negative organisms, where two regulatory components are involved: the transcriptional activator protein (black rectangle) and the AHL signaling molecule (oval) produced by the autoinducer synthase. The autoinducer accumulates in a cell density-dependent manner until a threshold level is reached. Then, the autoinducer binds to and activates the activator protein, inducing a change in gene expression. N-acyl homoserine lactone (AHL) is a signaling molecule used by the majority of gram-negative quorum-sensing systems. Adapted from de Kievit et al. with permission from the American Society for Microbiology, copyright 2000.3 (C) Illustrations of simple and complex switching in cell cycles. Left: the basic cell cycle in early embryos comprising nuclear and cell division (M phase) promoted by Cdk1, after Cdk2-directed DNA replication (S phase). Right: in adult tissues, the cell cycle contains a gap period (G1 phase) during which the activity of CDKs and other components are controlled by various positive and negative signals. Reprinted by permission from Macmillan Publishers Ltd: Nature,4 copyright 2004. (D–E) Studying switching with microfluidics. (D) Bright field microphotographs of plugs with clotted and unclotted human pooled blood plasma flowing through a microfluidic device (black arrows show direction of fluid flow). The fraction of plugs that clot increased from 0 to 1 as the concentration of thrombin increased from 0.0 to 0.6 nM. Adapted from Biophys. J.,5 copyright 2008, with permission from Elsevier. (E) Photograph of a 96-well plate showing detection of small increases in enzyme concentration with multiple threshold concentrations and visual readout. When the concentration of the enzyme acetylcholinesterase exceeds the concentration of the inhibitor, the enzyme hydrolyzes acetylthiocholine to give thiocholine, which results in a clear mixture. Reprinted with permission from Huynh et al.2 Copyright 2013 American Chemical Society. (F–G) Future avenues for microfluidic research related to switching. (F) Drawing of two examples of regulatory networks in yeast. Binding of a regulator (blue circle) to a gene promoter (red rectangle) is represented by a solid arrow, and genes encoding regulators are connected to their respective regulators by dashed arrows. From Lee et al.6 Reprinted with permission from AAAS. (G) Diagrams of a DNA circuit that computes either OR or AND depending on the initial concentration of the threshold. The graphs show results from kinetic experiments with digital logic gates, where input DNA strands were at 0.1× (0, logic OFF) or 0.9× (1, logic ON), where 1× = 100 nM. From Qian et al.7 Reprinted with permission from AAAS. | ||
The switching inherent in nature can be used to design artificial systems to detect the presence of target molecules or to control processes of reactions. Elaborate systems such as DNA circuits utilize simple reversible strand displacement reactions to build sophisticated digital logic circuits, where multiple “0” and “1” logic gates are integrated into a scaled-up cascade.7 This example fits within the recent field of synthetic biology, which seeks to modify or mimic biological systems to fulfill a particular purpose based on the construction of simple devices, such as transcription-based oscillators and switches.12 Rapid advances are being made in the field of synthetic chemical reaction networks that show dramatic examples of switching and other dynamic behaviors.13–15
Microfluidics and lab-on-a-chip technologies offer unique ways to study switching systems by precisely controlling microenvironments and addressing spatiotemporal effects with high resolution and throughput. For instance, microfluidics has been used to study the mechanisms of switching in blood clotting, as it affords precise control in the timing of reactions, mixing rates, and surface chemistry.5 Unlike bulk systems, where many experiments must be performed to determine the threshold concentration, microfluidic systems accelerate the search by enabling multiple reactions in parallel. When a threshold is multidimensional—i.e., when multiple chemical signals are interconnected in a complicated manner—microfluidics can capture the potential interactions between multiple components. Furthermore, switching can be used to engineer detection systems based on analog-to-digital conversion. For instance, an analog chemical signal can be translated into a series of digital yes-or-no bits to indicate whether biomarkers in a sample exceed known threshold concentrations, enabling microfluidic devices to perform measurements of clinically relevant changes in concentration of biomarkers.2 Furthermore, time control provided by microfluidics can enable “digitizing” reactions in time, instead of space: in one example, this approach was used to construct a chemical amplification network.16 However, microfluidic technology has a number of limitations as well. These include technical limitations, e.g., given the large surface-to-volume ratio in microfluidic systems, surface absorbance or evaporation may lead to inconsistent or inaccurate results when threshold systems are studied. At a higher level, microfluidic devices often struggle to capture the underlying realism and complexity of natural systems in vivo or at the cellular level. While microfluidics provides new avenues to accelerate in vitro or simple biological systems, it must be further developed in order to be applied widely in the study of complicated biological systems. Even so, the community is making progress in dealing with complexity, as shown in the emergence of organ-on-a-chip biomimetic technology.17
“Digital” as single entities
A second area of research within “digital” biology and chemistry is the analysis or detection of single biological entities such as molecules, cells, and organisms in isolation. In this context, spatial compartmentalization plays an important role and involves partitioning a solution or suspension of entities (such as cells) into different subunits. Poisson statistics is often important in this aspect and indicates that low ratios of biological entities to reaction compartments allow single entities to be captured per compartment. Microfluidics is a key approach for creating and manipulating such miniaturized fluidic compartments that contain single biological entities. We first present several examples of microfluidic technologies suitable for compartmentalization (Fig. 3) and follow with a discussion of the insights gained from these approaches. | ||
| Fig. 3 Microfluidic technologies for creating and manipulating fluidic compartments to study single biological or chemical entities. (A) Microphotograph of monodisperse water-in-oil emulsions for the encapsulation of single beads (top) and cells (bottom) used in droplet microfluidics. Adapted from Edd et al.18 (B) Schematic of elastomeric valve-based microfluidic large scale integration, used to divide a fluid stream (blue) into compartments by using valves (red). From Ottesen et al.19 Reprinted with permission from AAAS. (C) Photographs of SlipChip, in which wells in top and bottom plates overlap to form a continuous fluidic path (top). After slipping, the fluidic path is broken and fluid volumes are formed in each compartment (bottom). Adapted from Shen et al.20 (D) Illustration of beads loaded in femtoliter-sized wells. Beads settle onto the array of wells, after which a stream of sealing oil seals and isolates single beads inside microwells. Adapted from Kan et al.21 (E) Drawing showing formation of droplet compartments on hydrophilic-in-hydrophobic patterns by exchanging an aqueous solution with oil. Adapted from Sakakihara et al.22 (F) Composite fluorescence micrograph of antibody arrays obtained from microengraving single antibody-secreting cells. This approach was used to find single cells that produce monoclonal antibodies that recognize epitopes shared between two subtypes of a hepatitis B surface antigen. Reprinted by permission from Macmillan Publishers Ltd: Nat. Protoc.,23 copyright 2009. (G) Fluorescent micrograph of a spindle in a droplet in a microfluidic system that created cell-like compartments on a device. From Good et al.24 Reprinted with permission from AAAS. (H) Bright field and fluorescent microphotographs of bacteria in microfluidic wells, showing quorum sensing by a single cell. Bright field images (left) show the bacterium after 0 (top) and 17 h (bottom). Fluorescent images (right) show the activation of quorum sensing by a fluorescent reporter for gene expression of the gene LasB. Adapted from Boedicker et al.25 | ||
One example of a microfluidic technology used to study single entities is droplet microfluidics, which produces monodisperse droplets of aqueous reagents in the femto- to microliter range in an oil stream within microchannels. Miniaturization of chemical reactions by compartmentalization provides rapid mixing of reagents, as well as high control of interfacial properties and reaction timing.26 These droplets can be generated and manipulated at kilohertz rates and provide a valuable tool for studying compartments in high-throughput applications. Molecules, cells, organelles, and organisms, such as C. elegans, have already been encapsulated as single entities using this technology.27Fig. 3A gives an example of how intrinsic limitations set by Poisson statistics can be overcome by self-organizing high-density suspensions of cells in high-aspect ratio microchannels before encapsulation in droplets.18 Microfluidic large scale integration (LSI) is a technology broadly used for studying single cells and molecules. It relies on pneumatic valves that are monolithically fabricated in the silicone elastomer polydimethylsiloxane (PDMS) by using multilayer soft lithography.19 Compartmentalization of a sample is achieved by using a combinatorial array of binary valve patterns that, when closed, partition a network of microfluidic channels into compartments. After screening, any compartment can be selectively recovered from the device for further investigation. Another example includes SlipChip, a microfluidic device that can be used to confine single entities such as single molecules in compartments in a single step. The SlipChip consists of two plates that move relative to one another to form microfluidic channels and isolate microwells by creating and breaking fluidic paths depending on the position of one plate relative to the other.20 For this microfluidic format, no pumps or valves are required, making it suitable for limited-resource settings. The isolation and sealing of magnetic microbeads in femtoliter-sized reaction chambers is another technology that was successfully explored for single-molecule studies and detection.21 In this approach, microbeads are first used to capture single molecules. The beads are loaded and sealed in arrays of femtoliter-sized wells tailored to capture and seal one bead each. As such, reaction products generated by the single molecules are confined to very small volumes, ensuring a high local concentration of reaction products that can be detected with standard optical techniques. This approach was demonstrated on etched optical fiber bundles that were sealed with a gasket28 and on devices consisting of microwell arrays that are addressed by sequential fluid flows in a microchannel.21 Compartmentalization for single-molecule studies was also achieved by flowing an aqueous solution over hydrophilic-in-hydrophobic micropatterns and subsequently exchanging the aqueous solution with oil.22 As such, the hydrophilic micropatterns retain the aqueous solution as dome-shaped droplets while the aqueous solution in between the hydrophilic patterns is washed away, thereby forming many compartments simultaneously.
With the use of microfluidic compartmentalization, single molecules, organelles, cells, and organisms can be studied one by one in isolation to reveal information that would otherwise be invisible from ensemble measurements on seemingly homogeneous populations. Furthermore, analysis of biological entities at the single-entity level allows functional single entities to be found and recovered, as was demonstrated for screening and retrieving individual antibody-secreting cells by a lithographic process called microengraving.23 Compartmentalization with microfluidics also offers perspectives for reconstructing and understanding intracellular processes. For instance, a microfluidic system that encapsulated cytoplasm from Xenopus eggs and embryos inside droplets of defined sizes demonstrated how cytoplasmic volume modulates spindle size during embryogenesis.24 Furthermore, co-compartmentalization allows the study of interactions between biological entities at the single-entity level, as was achieved for virus-bacteria interactions when microfluidic digital PCR was used to examine the association between viral gene markers and single bacteria.29
The study of naturally switching systems at the single-entity level in compartments offers a promising tool for improving our understanding of the heterogeneity of populations that exhibit switching. A study employing single-cell compartmentalization and subsequent qRT-PCR on a Fluidigm Biomark chip, for example, found bimodal expression of key immune genes in mouse bone marrow-derived dendritic cells in response to lipopolysaccharide, even for genes that are highly expressed at the population average.30 In addition, it was shown that confinement of single bacteria in small volumes can initiate quorum sensing and quorum sensing-dependent growth, while the initiation of quorum sensing varies highly within the same clonal population.25 This demonstrates the potential of confinement to be used for understanding and controlling autocrine signaling at the single-cell level for better understanding of such pathways. Finally, isolation and confinement of single biological entities in compartments also allow reaction products to be confined and achieve high local concentrations that can result in sensitive detection of single molecules and cells, as discussed in more detail in the following section.
Apart from microfluidic compartmentalization, other methods have been reported that allow single biological entities to be studied without the need for compartmentalization, such as optical super-resolution microscopy and combinatorial labeling31 and flow cytometric analysis (FACS)32 for single-cell studies. While the advantages of microfluidic compartmentalization are apparent when dealing with secreted signals, compartmentalization can also be a disadvantage: for instance, biomolecules and cells function within contexts of networks, cells, and organisms, but these complex interactions may not always be captured by compartmentalization. One of the challenges for microfluidics in digital biology and chemistry is to start capturing and maintaining biological context and complexity, such as by controlling interactions between cells.
Using compartmentalization of single entities to induce switching in non-switching systems and enable sensitive detection
Compartmentalization of single entities can also be used to induce switching in systems that do not naturally switch. Here, we focus on one specific application of this strategy: quantifying molecules and cells (Fig. 4). Examples include ELISA, PCR, and other amplification reactions that are commonly used to measure the concentration of proteins, nucleic acids, or other entities. When used in the traditional “bulk” format (e.g., in tubes or microtiter plates), quantification is achieved by monitoring the kinetics of the amplification reaction and comparing it with standard concentrations, but no critical threshold in input concentration changes the output drastically from “off” to “on”. However, if single entities are compartmentalized prior to amplification, each compartment will produce a clear “on” or “off” signal after amplification depending on whether a target molecule was or was not present in each compartment. Quantification of the target molecules present in the starting solution can thus be performed by simply counting the fraction of compartments turning “on” after the amplification reaction (endpoint detection), with no requirement to monitor the reaction kinetics.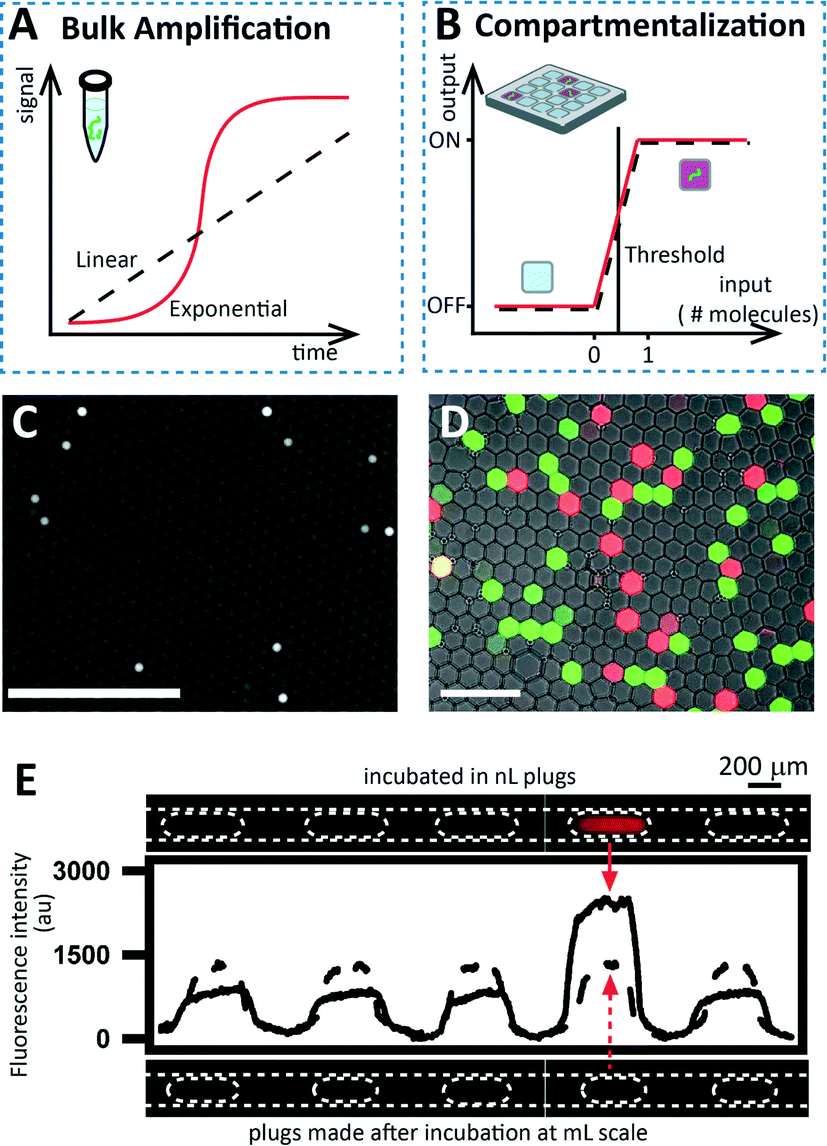 | ||
| Fig. 4 Using compartmentalization to induce switching in non-switching systems. (A) Graph showing that two amplification reactions used to quantify the concentration of target molecules do not exhibit switching when performed in bulk. One reaction (dotted line) represents linear kinetics (e.g., in ELISA), while the other (red curve) represents exponential kinetics (e.g., PCR). (B) Graph showing how compartmentalization transforms the systems described in (A) into switching systems. If a compartment contains at least one target molecule (input), the signal will turn “on” in the endpoint detection. If the compartment does not contain a molecule, no amplification will happen and the reaction will stay “off”. (C) Cropped photograph of digital ELISA, obtained by confining beads before enzymatic reaction and readout. Reprinted by permission from Macmillan Publishers Ltd: Nat. Biotechnol.,28 copyright 2010. (D) A cropped photograph of digital PCR in microfluidic droplets, performed with two different Taqman probes for two-plex detection. Adapted from Pekin et al.34 (E) Linescans and microphotographs showing detection of bacteria by confinement in nanoliter plugs. The upper microphotograph shows that only the plug containing one cell showed a positive signal after 2.8 hours. Adapted from Boedicker et al.37 | ||
A well-known example of switching induced by compartmentalization is digital ELISA,28 for which a platform is commercially available from Quanterix. In this case, the immunocomplex is formed on the surface of magnetic beads in solution and labeled with an enzymatic reporter generating a fluorescent product. Single beads are placed and isolated in single wells before the enzymatic reaction. Since the effective volume of liquid in each well is on the order of femtoliters, the signal of a single molecule is detectable in a few minutes. This has allowed limits of detection that are much lower than the conventional ELISA.
Compartmentalization has also been used to enhance the properties of DNA/RNA amplification reactions, such as PCR and isothermal chemistries. This approach to PCR was demonstrated when the DNA template was serially diluted and analyzed by conventional PCR.33 For each dilution, the number of replicates showing an “on” signal after amplification was monitored, allowing an estimation of the starting concentration without the need for an added standard. The potential of this approach was enhanced by the use of microfluidics, which enabled a wider dynamic range through the use of a larger number of volumes in parallel. Strategies have been applied in various formats, including microwells,19 droplets,34 and the SlipChip,20,35,36 to name a few, and several platforms for digital, single-molecule nucleic acid amplification, such as those available from Fluidigm Corporation, Bio-Rad Laboratories, and RainDance Technologies, are commercially available.
The use of compartmentalization to induce switching was also demonstrated for the detection of single bacteria.37 In this approach, bacteria were compartmentalized into nanoliter plugs containing alamarBlue, a fluorescent viability indicator. When live cells are incubated in the presence of this indicator, electron receptors commonly produced during metabolic activity reduce the active ingredient (resazurin), producing a fluorescent signal. Performing this assay in nanoliter plugs allowed compartmentalization of the bacteria, and bacteria could be quantified by counting the plugs that were switched “on.” In addition, the effective concentration was increased in the plugs containing at least one cell, so the time needed to generate a detectable signal was reduced compared to the bulk experiment.
Digitizing these non-switching systems offers significant advantages in terms of assay design, simplicity, and robustness because the yes-or-no format makes reaction kinetics less critical. Since endpoint detection can be used, as opposed to tracking reactions in real-time, quantitative information can be obtained with qualitative measurements. This ability makes digital assays more robust to small fluctuations in reaction conditions, as compared to bulk equivalents based on kinetic measurements (Fig. 5A–B). Reaction conditions can be affected by a large number of parameters, including temperature, pH, concentration of reagents, concentration of target molecules, and presence of inhibitors.
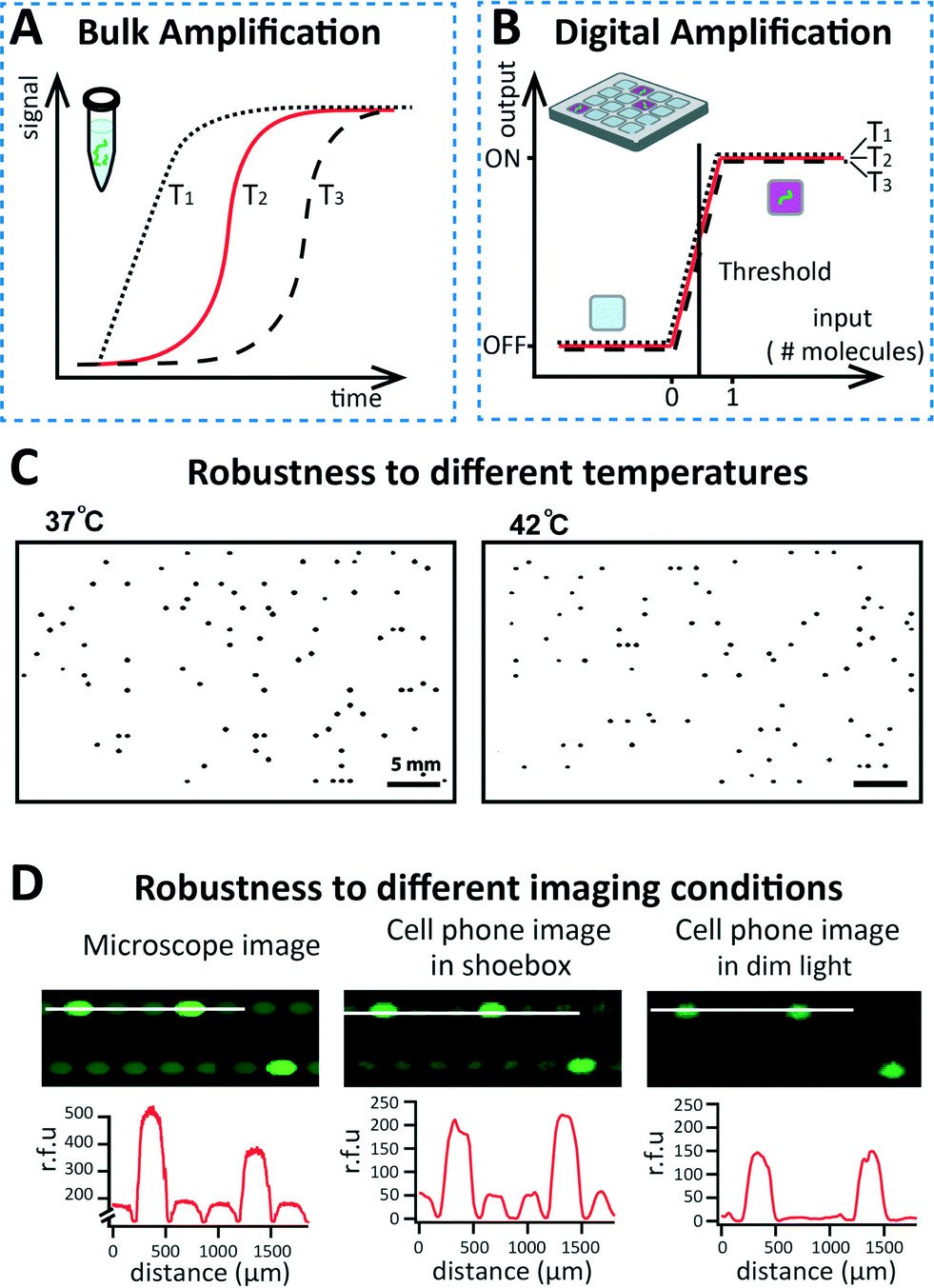 | ||
| Fig. 5 Robustness of digital detection approaches. (A) Schematic drawing showing hypothetical traces of three isothermal amplification reactions performed in bulk using the same concentration of target molecules. Differences in temperature affect the kinetics, giving rise to differences in the reaction profiles. (B) Schematic drawing showing that if the same reactions from (A) are performed in a digital format, they show robustness to temperature fluctuations, giving similar results. (C) Representative fluorescence images of MRSA gDNA amplification by RPA on SlipChip, performed at two different temperatures (37 °C and 42 °C), show comparable counts. Images were inverted and processed using a Gaussian blur and threshold to enhance contrast. Reprinted with permission from Shen et al.35 Copyright 2011 American Chemical Society. (D) Images of digital LAMP (top row) performed on SlipChip and imaged under three different conditions: (i) using a microscope, (ii) using a cell phone in a dark environment, and (iii) using a cell phone in dim light. The intensity profiles (bottom row) are comparable, showing robustness to different imaging conditions. Reprinted with permission from Selck et al.36 Copyright 2013 American Chemical Society. | ||
For known amplification chemistries, increased robustness by digitization has already resulted in more sensitive quantitative measurements. The effect of fluctuations in temperature were evaluated in the case of digital RPA35 (Fig. 5C) and digital LAMP36 on SlipChip. The total number of positive wells was not significantly affected even with temperature changes of up to 5 °C for RPA (Fig. 5C) and 6 °C for LAMP. Moreover, the use of yes/no measurements allowed simplified detection and increased robustness. Digital LAMP reactions performed on SlipChip were detected with a cell phone camera in a dark box, and even in dim lighting (see Fig. 5D). In addition, images were automatically processed by custom software, giving results comparable to the standard detection performed with a microscope (Video S1†).36 This improved robustness of digital formats opens the road to using new chemistries for quantitative measurements. Amplification strategies that would typically not be considered for this purpose due to their kinetics (e.g., noisy chemistries lacking a smooth input/output relationship) can be harnessed for quantitative measurements.
Outlook
Here we have discussed two areas of research in digital biology and chemistry that we find particularly interesting: the study of systems that switch between discrete states, and the study of single biological entities. We have illustrated strong bidirectional connections between these areas and microfluidic technologies: microfluidics offers a number of tools that enable new advances in these areas, and conversely, digital biology and chemistry enable new capabilities for lab-on-a-chip approaches, such as in microanalytical systems.We expect that these connections will continue to strengthen in the future. One area that is ripe for a broader investigation is the study of switching in synthetic chemical reaction networks, many of which involve strong degrees of amplification that would enable single-molecule detection and characterization.7,13–15 Microfluidics can be used to advance fundamental understanding of the dynamics of such networks, and can in turn benefit from dramatically improved detection capabilities likely to result from such work. “Digitizing” these networks in time16 in addition to compartmentalizing in space is an opportunity likely to lead to systems that are certainly interesting, and are possibly useful.
Several areas are being actively investigated due to the confluence of well-posed biological questions and established technological capabilities. The mammalian immune system presents a fertile area of research: it shows incredible heterogeneity, e.g., in the T cell and B cell repertoire, and also exhibits complex switching patterns governed by a multi-dimensional set of signals, such as in many aspects of development in the immune system. Analysis of DNA and RNA enabled by high-throughput sequencing technologies is matched well to the complexity of these problems.
Not all of the questions in digital biology and chemistry will be addressable by analysis of nucleic acids, however. For example, single-cell and single-molecule level analysis of proteins, carbohydrates, lipids, small molecules, and other cellular and molecular components will require further technological development to enable digital biology and chemistry. Emerging technologies for single-cell protein analysis are already making an impact. Moving these and other technologies from the “detection” mode (detecting and quantifying known molecules) to the “discovery” mode (identifying and quantifying unknown molecules) will further broaden the impact of digital biology and chemistry.
As digital biology and chemistry move from examining individual entities to understanding interactions among them, these disciplines will address fascinating questions in a number of fields of biology. We find particularly appealing the fields of development, immunology, and especially microbial communities, where complex switching dynamics and heterogeneity coexist with incredible networks of interactions among microbes and often include their hosts. These advances would require development of new generations of lab-on-a-chip systems and analytical methods.
Broadly speaking, while many exciting microfluidic tools for digital analysis already exist and many new ones will be developed, two challenges should be kept in mind: (i) integrating these technologies with upstream processing of samples and with downstream processing of information and (ii) making these tools massively distributed, both in terms of cost and ease of operation. We and others have been working to address these challenges. For example, Video S2† shows that a six-year-old child can make thousands of “digital” compartments on a SlipChip device, and another video36,38 shows how such devices can take advantage of the robustness of digital analysis to enable a five-year-old user to automatically quantify viral RNA molecules using a cell phone connected to a cloud server (Video S1†). Such widely distributed, simple-to-use quantitative digital platforms would substantially accelerate research in chemical and biological laboratories. Beyond research, these systems could expand access to medical diagnostics to homes and primary care clinics, positively impact agricultural biotechnology, and enable worldwide pandemic surveillance of emerging diseases.
Acknowledgements
We would like to thank the five-year-old volunteer for performing the demonstration in Video S1†, the six-year-old volunteer for performing the demonstration shown in Video S2†, and Liang Li of SlipChip Corp. for providing the chip used in this experiment. Disclosure: R.F.I. has a financial interest in SlipChip Corp.References
- K. Choi, A. H. C. Ng, R. Fobel and A. R. Wheeler, Annu. Rev. Anal. Chem., 2012, 5, 413–440 CrossRef CAS PubMed.
- T. Huynh, B. Sun, L. Li, K. P. Nichols, J. L. Koyner and R. F. Ismagilov, J. Am. Chem. Soc., 2013, 135, 14775–14783 CrossRef CAS PubMed.
- T. R. de Kievit and B. H. Iglewski, Infect. Immun., 2000, 68, 4839–4849 CrossRef CAS.
- J. Massagué, Nature, 2004, 432, 298–306 CrossRef PubMed.
- R. R. Pompano, H. W. Li and R. F. Ismagilov, Biophys. J., 2008, 95, 1531–1543 CrossRef CAS PubMed.
- T. I. Lee, N. J. Rinaldi, F. Robert, D. T. Odom, Z. Bar-Joseph, G. K. Gerber, N. M. Hannett, C. T. Harbison, C. M. Thompson, I. Simon, J. Zeitlinger, E. G. Jennings, H. L. Murray, D. B. Gordon, B. Ren, J. J. Wyrick, J.-B. Tagne, T. L. Volkert, E. Fraenkel, D. K. Gifford and R. A. Young, Science, 2002, 298, 799–804 CrossRef CAS PubMed.
- L. Qian and E. Winfree, Science, 2011, 332, 1196–1201 CrossRef CAS PubMed.
- M. A. Savageau, Nature, 1974, 252, 546–549 CrossRef CAS.
- H. H. Ng, Y. Zhang, B. Hendrich, C. A. Johnson, B. M. Turner, H. Erdjument-Bromage, P. Tempst, D. Reinberg and A. Bird, Nat. Genet., 1999, 23, 58–61 CAS.
- G. I. Evan and K. H. Vousden, Nature, 2001, 411, 342–348 CrossRef CAS PubMed.
- R. Medzhitov, Nature, 2007, 449, 819–826 CrossRef CAS PubMed.
- S. A. Benner and A. M. Sismour, Nat. Rev. Genet., 2005, 6, 533–543 CrossRef CAS PubMed.
- S. N. Semenov, A. J. Markvoort, W. B. L. Gevers, A. Piruska, T. F. A. de Greef and W. T. Huck, Biophys. J., 2013, 105, 1057–1066 CrossRef CAS PubMed.
- T. Bánsági, V. K. Vanag and I. R. Epstein, Science, 2011, 331, 1309–1312 CrossRef PubMed.
- X. Chen, N. Briggs, J. R. McLain and A. D. Ellington, Proc. Natl. Acad. Sci. U. S. A., 2013, 110, 5386–5391 CrossRef CAS PubMed.
- C. J. Gerdts, V. Tereshko, M. K. Yadav, I. Dementieva, F. Collart, A. Joachimiak, R. C. Stevens, P. Kuhn, A. Kossiakoff and R. F. Ismagilov, Angew. Chem., Int. Ed., 2006, 45, 8156–8160 CrossRef CAS PubMed.
- D. Huh, B. D. Matthews, A. Mammoto, M. Montoya-Zavala, H. Y. Hsin and D. E. Ingber, Science, 2010, 328, 1662–1668 CrossRef CAS PubMed.
- J. F. Edd, D. Di Carlo, K. J. Humphry, S. Koster, D. Irimia, D. A. Weitz and M. Toner, Lab Chip, 2008, 8, 1262–1264 RSC.
- E. A. Ottesen, J. W. Hong, S. R. Quake and J. R. Leadbetter, Science, 2006, 314, 1464–1467 CrossRef CAS PubMed.
- F. Shen, W. Du, J. E. Kreutz, A. Fok and R. F. Ismagilov, Lab Chip, 2010, 10, 2666–2672 RSC.
- C. W. Kan, A. J. Rivnak, T. G. Campbell, T. Piech, D. M. Rissin, M. Mosl, A. Peterca, H. P. Niederberger, K. A. Minnehan, P. P. Patel, E. P. Ferrell, R. E. Meyer, L. Chang, D. H. Wilson, D. R. Fournier and D. C. Duffy, Lab Chip, 2012, 12, 977–985 RSC.
- S. Sakakihara, S. Araki, R. Iino and H. Noji, Lab Chip, 2010, 10, 3355–3362 RSC.
- A. O. Ogunniyi, C. M. Story, E. Papa, E. Guillen and J. C. Love, Nat. Protoc., 2009, 4, 767–782 CrossRef CAS PubMed.
- M. C. Good, M. D. Vahey, A. Skandarajah, D. A. Fletcher and R. Heald, Science, 2013, 342, 856–860 CrossRef CAS PubMed.
- J. Q. Boedicker, M. E. Vincent and R. F. Ismagilov, Angew. Chem., Int. Ed., 2009, 48, 5908–5911 CrossRef CAS PubMed.
- H. Song, D. L. Chen and R. F. Ismagilov, Angew. Chem., Int. Ed., 2006, 45, 7336–7356 CrossRef CAS PubMed.
- M. T. Guo, A. Rotem, J. A. Heyman and D. A. Weitz, Lab Chip, 2012, 12, 2146–2155 RSC.
- D. M. Rissin, C. W. Kan, T. G. Campbell, S. C. Howes, D. R. Fournier, L. Song, T. Piech, P. P. Patel, L. Chang, A. J. Rivnak, E. P. Ferrell, J. D. Randall, G. K. Provuncher, D. R. Walt and D. C. Duffy, Nat. Biotechnol., 2010, 28, 595–599 CrossRef CAS PubMed.
- A. D. Tadmor, E. A. Ottesen, J. R. Leadbetter and R. Phillips, Science, 2011, 333, 58–62 CrossRef CAS PubMed.
- A. K. Shalek, R. Satija, X. Adiconis, R. S. Gertner, J. T. Gaublomme, R. Raychowdhury, S. Schwartz, N. Yosef, C. Malboeuf, D. Lu, J. J. Trombetta, D. Gennert, A. Gnirke, A. Goren, N. Hacohen, J. Z. Levin, H. Park and A. Regev, Nature, 2013, 498, 236–240 CrossRef CAS PubMed.
- E. Lubeck and L. Cai, Nat. Methods, 2012, 9, 743–748 CrossRef CAS PubMed.
- B. Bodenmiller, E. R. Zunder, R. Finck, T. J. Chen, E. S. Savig, R. V. Bruggner, E. F. Simonds, S. C. Bendall, K. Sachs, P. O. Krutzik and G. P. Nolan, Nat. Biotechnol., 2012, 30, 858–867 CrossRef CAS PubMed.
- P. J. Sykes, S. H. Neoh, M. J. Brisco, E. Hughes, J. Condon and A. A. Morley, Biotechniques, 1992, 13, 444–449 CAS.
- D. Pekin, Y. Skhiri, J.-C. Baret, D. Le Corre, L. Mazutis, C. Ben Salem, F. Millot, A. El Harrak, J. B. Hutchison, J. W. Larson, D. R. Link, P. Laurent-Puig, A. D. Griffiths and V. Taly, Lab Chip, 2011, 11, 2156–2166 RSC.
- F. Shen, E. K. Davydova, W. Du, J. E. Kreutz, O. Piepenburg and R. F. Ismagilov, Anal. Chem., 2011, 83, 3533–3540 CrossRef CAS PubMed.
- D. A. Selck, M. A. Karymov, B. Sun and R. F. Ismagilov, Anal. Chem., 2013, 85, 11129–11136 CrossRef CAS PubMed.
- J. Q. Boedicker, L. Li, T. R. Kline and R. F. Ismagilov, Lab Chip, 2008, 8, 1265–1272 RSC.
- http://pubs.acs.org/doi/suppl/10.1021/ac4030413/suppl_file/ac4030413_si_002.mp4, Accessed February 24, 2014.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c4lc00248b |
| This journal is © The Royal Society of Chemistry 2014 |