
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceStable end-sealed DNA as robust nano-rulers for in vivo single-molecule fluorescence†
A.
Plochowietz
*a,
A. H.
El-Sagheer
bc,
T.
Brown
 b and
A. N.
Kapanidis
*a
b and
A. N.
Kapanidis
*a
aDepartment of Physics, University of Oxford, Clarendon Laboratory, Parks Road, Oxford, OX1 3PU, UK. E-mail: anne.plochowietz@gmail.com; a.kapanidis1@physics.ox.ac.uk
bDepartment of Chemistry, University of Oxford, Chemistry Research Laboratory, 12 Mansfield Road, Oxford, OX1 3TA, UK
cChemistry Branch, Department of Chemistry, Faculty of Petroleum and Mining Engineering, Suez University, Suez 43721, Egypt
First published on 23rd March 2016
Abstract
Single-molecule fluorescence and Förster resonance energy transfer (smFRET) are important tools for studying molecular heterogeneity, cellular organization, and protein structure in living cells. However, in vivo smFRET studies are still very challenging, and a standardized approach for robust in vivo smFRET measurements is still missing. Here, we synthesized protected DNAs with chemically linked ends as robust in vivo nano-rulers. We efficiently internalized doubly-labeled end-sealed DNA standards into live bacteria using electroporation and obtained stable and long-lasting smFRET signatures. Single-molecule fluorescence signals could be extended to ∼1 min by studying multi-fluorophore DNA standards. The high stability of protected DNA standards offers a general approach to evaluate single-molecule fluorescence and FRET signals, autofluorescence background, and fluorophore density, and hence, quality check the workflow for studying single-molecule trajectories and conformational dynamics of biomolecules in vivo.
Introduction
Single-molecule fluorescence and single-molecule Förster resonance energy transfer (smFRET) studies have advanced substantially our understanding of molecular and cellular processes over the last two decades.1–3 Single-molecule fluorescence is increasingly employed in vivo to study gene expression stochasticity and spatial organization of biomolecules in the natural cellular environment, whereas smFRET is well suited for studying protein structure and dynamics both in vitro and in living cells.4,5 FRET relies on the non-radiative energy transfer from a donor fluorophore (D) to a complementary acceptor fluorophore (A) present in close proximity (2–10 nm).6–8In vitro smFRET has been used extensively to study many processes including nucleic acid and protein folding,9,10 and conformational changes of large protein complexes;11–15 these studies enabled structure-function single-molecule analysis and uncovered mechanistically relevant molecular heterogeneities.Despite the extensive use of smFRET in vitro, live-cell smFRET studies are still challenging, mainly due to the difficulty of site-specific FRET dye-pair labeling of biomolecules in living cells.16 Fluorescent proteins (FPs) are often employed as in vivo FRET dye-pairs, but their photophysical properties (blinking, poor photostability, low brightness) prevent their use in single-molecule FRET studies.17,18 Further, labeling strategies using FPs (∼100-fold larger than organic dyes) are limited to protein end-labeling.5
In contrast, organic dyes are much better suited for smFRET; however, they have to be introduced into live cells by in vivo specific protein labeling via polypeptide tags (SNAP, HALO, or TMP tags19–21) or via unnatural amino acids;22 alternatively, delivery can rely on internalization of in vitro organic-dye labeled proteins into live cells. The latter strategy was used in a handful of smFRET studies in live prokaryotic23 and eukaryotic24,25 cells. In one of these approaches, we used electroporation to internalize doubly-labeled DNAs and DNA-binding proteins into live bacteria23,26 and characterized organic dyes for their use in in vivo FRET studies.27
To characterize in vivo FRET measurements, we previously used blunt-ended 45-bp double-stranded DNA with different donor–acceptor distances to monitor low-, intermediate-, and high-FRET signals inside single cells. In those studies, we observed decreased FRET for some of the internalized DNA compared to in vitro measurements,23,27 and attributed this shift mainly to in vivo DNA degradation by endonucleases that recognize blunt DNA ends and digest DNA.28
The absence of robust DNA standards that report on FRET, degradation processes, and cellular autofluorescence has slowed down the implementation of single-molecule fluorescence and FRET studies in living cells. Here, we address this limitation by introducing doubly-labeled protected DNA FRET standards and multi-fluorophore protected DNAs, in which both DNA ends are chemically linked using click chemistry (Scheme 1, ESI†) to prevent DNA degradation inside live E. coli. The protection of oligonucleotide termini was previously demonstrated for cyclic dumbbell DNA duplexes.29,30 Here, the CuAAC reaction was chosen for three reasons: (i) its high efficiency in aqueous media, (ii) its orthogonality with the functional groups of DNA and the primary amino groups used as attachment points for fluorescent dyes, and (iii) the simplicity of introducing the alkyne and azide functions into oligonucleotide strands.
 | ||
| Scheme 1 Templated click ligation to synthesize end-sealed DNA duplexes; fluorophores are shown for intermediate-FRET (P18), high-FRET (P8), and multi-fluorophore DNA standards (P(Cy3B)4). | ||
Results
Our protected DNA FRET standards were initially characterized in vitro and internalized into live E. coli using electroporation. We employed alternating laser excitation (ALEX, ref. 31 and 32) in vivo to identify donor–acceptor molecules and show that their FRET values agree very well with our in vitro measurements. We also combined smFRET measurements with single-particle tracking and obtained stable and long-lasting smFRET trajectories (∼10 s), and multi-fluorophore DNA trajectories (∼1 min), showing that the protected DNAs are well suited to monitor smFRET levels in living cells.We synthesized doubly-labeled 45-bp protected DNAs with different dye spacing corresponding to intermediate-FRET efficiencies (18 bp spacing, hereafter P18), and high-FRET efficiencies (8 bp spacing, hereafter P8; Scheme 1, ESI†). We used the FRET pair Cy3B/Atto647N, which we previously showed to perform well in single-cell FRET studies.27
To characterize the stability of the protected DNA FRET standards in vitro and test for any effects of their exposure to electroporation conditions (as tested in the electroporation cuvette but in the absence of cells), we used confocal ALEX microscopy (Experimental section). Both the fluorescence intensity time-traces and their autocorrelation function of electroporated protected DNAs (ACF; ESI†) showed the typical burst duration (∼1–2 ms) expected for a DNA of their size, and indicated the presence of a single diffusing species both before and after electroporation (Fig. S1†). This was in contrast to unprotected, blunt-ended DNA FRET standards, for which DNA aggregated during electroporation (Fig. S2;† 20–30 ms burst length); this aggregation was overcome by adding 1 mM EDTA to blunt-ended DNAs before electroporation (Fig. S2†), likely due to EDTA chelating Al3+-ions released from the electroporation cuvette.33
Sorting the fluorescence bursts in 2D-histograms of FRET (E) and probe stoichiometry (S, a fluorescence ratio that reports on molecular stoichiometry, ESI†), we observed a single FRET species both for electroporated P18 (E ∼ 0.42) and electroporated P8 (E ∼ 0.89) (Fig. 1). The excellent agreement of ES-histograms for the FRET standards before and after electroporation for six different electroporation voltages (0.8–1.8 kV, Fig. S3†), as well as the absence of free dye26 (Fig. S4†) make the protected DNAs well suited for internalization into live bacteria. FRET and stoichiometry values were corrected for cross-talk contribution and different detector efficiencies at each emission channel (γ-factor). Technical detail are given in the ESI,† the contribution of the different correction terms to the ES-histograms for in vitro measurements are shown in Fig. S5,† and the estimation of the γ-factor is shown in Fig. S6.†
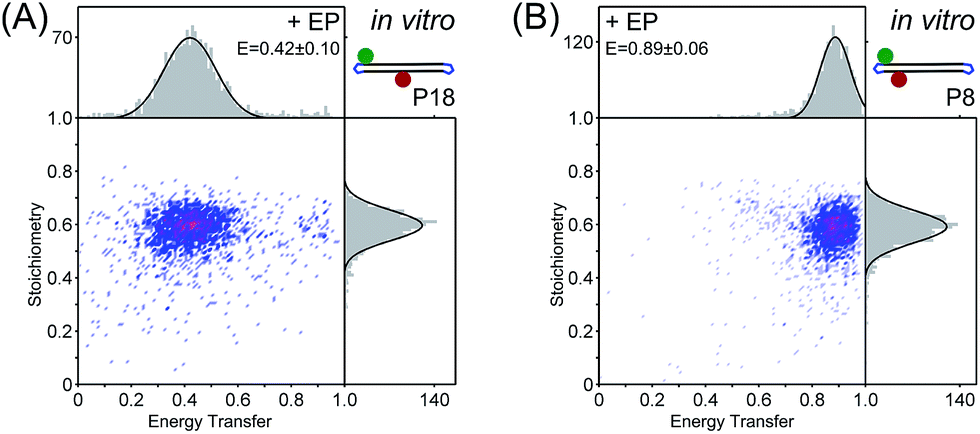 | ||
| Fig. 1 In vitro characterization of (A) protected intermediate-FRET DNA (P18) and (B) protected high-FRET DNA (P8) using confocal ALEX microscopy. FRET-stoichiometry histograms show single intermediate- and high-FRET species after electroporation with single Gaussian fits centered at E = 0.42 ± 0.10 and E = 0.89 ± 0.06 (±standard deviation), respectively. E values are corrected for cross-talk contributions and different detector efficiencies at each emission channel,32 see ESI and Fig. S5.† | ||
To evaluate the performance of the protected DNA standards in vivo, we electroporated them into live E. coli, recovered electroporated cells, and removed non-internalized DNAs by washing before imaging (Experimental section). We applied an initial electric field of 14 kV cm−1, which maintained ∼70% cell viability27 and resulted in a cellular uptake of up to 8 DNA molecules per cell (median: ∼1 molecule per cell). We imaged cells on agarose pads by an inverted wide-field fluorescence microscope using HILO illumination34 combined with ALEX (Experimental section).
We sorted species (Fig. 2A) into FRET molecules (which contain both an emitting donor and an emitting acceptor), donor-only molecules, and acceptor-only molecules. Single-molecule localization of FRET molecules was first performed using the FRET channel (i.e., donor-excitation/acceptor emission channel; DA channel), where there is little interference from donor-only, acceptor-only species, and autofluorescence; we then mapped the DA signal to the corresponding signal in the donor-excitation/donor emission channel (DD channel), and linked these signals to the acceptor-excitation/acceptor emission channel (AA channel; ESI†). We only included molecules with >400 photons per frame (for DD + DA) above the cellular autofluorescence to ensure accurate fitting of single molecule images in the respective channels (Fig. S7, ESI†); the 2D Gaussian fitting of the images accounted for the different cellular autofluorescence in the respective fluorescence channels.
 | ||
| Fig. 2 In vivo characterization of protected DNA FRET standards. (A) Electroporation of protected high FRET DNA standards into live E. coli and observation of smFRET signals using an alternating laser excitation scheme (ALEX) monitoring D-only (DD channel), A-only (AA channel), and smFRET (DA channel) molecules. (B–C) Accurate FRET-stoichiometry histograms of P18 and P8 standards; the main FRET species are centered at E = 0.40 ± 0.18 and at E = 0.87 ± 0.05, respectively. N denotes the number of selected molecules P18: ∼45%, and P8: ∼30% of all initial localizations, 3 independent data sets are combined (∼1000 cells each, 0–2 FRET molecules per cell) and example cell images are shown as a composite of DD- and DA-channel (overlay) such that high-FRET, intermediate-FRET, and D-only DNA molecules appear red, yellow, and green, respectively. | ||
The ES histograms for the selected species were also corrected for cross-talk and γ-factor contributions (Fig. S8†) and showed a single FRET species for P18 (E = 0.40 ± 0.18; Fig. 2B), and for P8 (E = 0.87 ± 0.05; Fig. 2C). The mean FRET efficiencies obtained in vivo were in excellent agreement with in vitro FRET values (0.42 and 0.89, respectively) and did not show a shift towards smaller FRET values as previously seen for immobile blunt-ended DNA.23 The increased width of the fitted Gaussian distribution was due to the in vivo smFRET signals being noisier than in vitro signals, which we mainly attribute to effects of molecular motion, cellular autofluorescence, and nearby molecules.
To test our ability to perform extended observations using protected DNAs, we tracked single FRET molecules in single cells while monitoring their FRET signatures in green continuous-wave mode (ESI†). We tracked smFRET molecules for >10 s and obtained stable and long-lasting smFRET time-traces (Fig. 3A and B). The time-traces show different levels of noise due to molecular motion and cellular autofluorescence. The smFRET observation time is mainly limited by photobleaching, but can be extended to the min-timescale using time-lapse imaging.
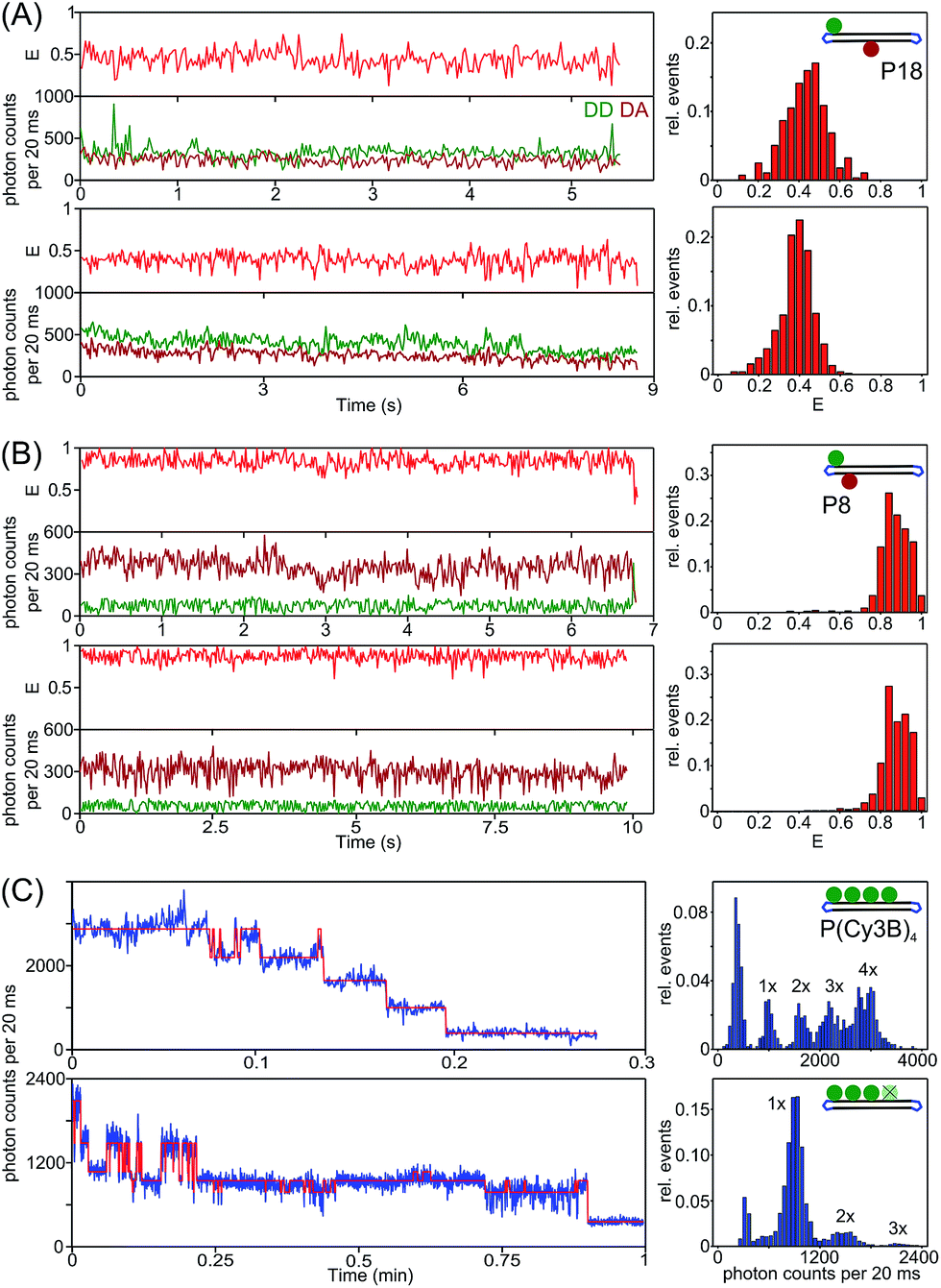 | ||
| Fig. 3 Long-lasting observation of protected DNA standards using single-particle tracking. (A–B) Accurate smFRET time-traces and FRET-histograms showing E-values (red), DD-intensity (dark green), and DA-intensity (dark red) of single protected intermediate and high DNA FRET molecules, respectively. These FRET time-traces showed anti-correlated DD and DA-signals, a hallmark of single-molecule FRET (e.g. B: top time-trace) and spikes in DD-signal due to nearby D-only molecules (e.g. A: top time-trace). More smFRET showing donor and acceptor photobleaching events are shown in Fig. S9.† (C) Single-molecule fluorescence time-traces of multi-fluorophore protected DNA showing distinct photobleaching events and fluorophore blinking (blue: raw data, red: HMM fit, ESI†). Single-molecule intensity histograms show 3–4 Cy3B-labels per single DNA molecule. | ||
To extend the single-molecule fluorescence observation of internalized DNAs, we also studied a protected multi-fluorophore DNA standard labeled with 4 Cy3B fluorophores (P(Cy3B)4, Scheme 1). Long-lasting (>10 s) single-molecule fluorescence time-traces were picked and Hidden Markov Modeling analysis was applied to elucidate single photobleaching and blinking events. We observed up to 4 distinct photobleaching steps and fluorescence signal from a single DNA molecule for >50 s (Fig. 3C). The fluorescence intensity per Cy3B fluorophore was estimated to ∼30![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 photons per s, which could increase the temporal resolution by 2–3 fold, and extend single-molecule observation to a few minutes.
000 photons per s, which could increase the temporal resolution by 2–3 fold, and extend single-molecule observation to a few minutes.
Conclusions
Our results establish that protected DNAs can serve as excellent standards for in vivo single-molecule fluorescence due their efficient internalization into E. coli and their long-lived stability. Interestingly, our results show that the cellular environment does not dramatically affect the fluorophore properties (at least when fluorophores are photoactive) and corrected FRET efficiencies of the standards (no significant DNA degradation), and that in vitro and in vivo smFRET efficiencies agreed remarkably well, despite being acquired in different media (cellular cytoplasm vs. an aqueous buffer). In addition, we were able to obtain stable smFRET time-traces for ∼10 s and long-lasting multi-fluorophore single-molecule fluorescence time-traces of >50 s in the bacterial cytoplasm. These results make the protected DNA standards ideal for robust single-molecule fluorescence observation and help to implement smFRET in living cells.The in vivo use of DNA standards can be further improved. Due to the wide range of internalized molecules per cell (0–8, median: 1 molecule per cell), single-particle tracking was impeded by nearby molecules (ideally: ∼1–3 molecules per cell, 1 mol μm−2, ref. 35). This “crowding” scenario, along with higher autofluorescence (especially in the DD channel) also skewed smFRET signals (e.g., due to DD-signal spikes, Fig. 3A), since the single-molecule localization routine relies on assuming that only a single fluorophore contributes to the fluorescence signal. Thus, performing smFRET in the 650–800 nm range (e.g. using Cy5/Cy7, and Atto680/Atto740, ref. 18), where autofluorescence is minimized, will offer a “cleaner” spectral window for smFRET.
The protected DNAs should be used as reference “nano-rulers” to calibrate and check the quality of in vivo smFRET measurements, report on cellular autofluorescence background, and help optimize the fluorophore density for single-molecule observation in living cells. The stable end-sealed DNAs are versatile and can easily be modified to become standards for co-localization studies, multi-color experiments beyond FRET, or reporters on complex stoichiometries such as the P(Cy3B)4 standard. Finally, protected DNA substrates with modified functional groups and varying DNA sequences could be employed to study specific DNA–protein interactions and their spatial distribution within the living cell. These dsDNA can act as ‘decoys’ (e.g. copies of transcription factor sites) and effectively compete with chromosomal sites and alter gene expression.
Experimental
DNA sequences are shown 5′ to 3′:S1: ![[T with combining low line]](https://www.rsc.org/images/entities/char_0054_0332.gif) AAATCTAAAGTAACATAAGGTAACATAACGTAAGCTCATTCGCG
AAATCTAAAGTAACATAAGGTAACATAACGTAAGCTCATTCGCG
S2: CGCGAATGAGCTTACGTTATGTTACCTTATGTTACTTAGATTTA
S3: CGCGAATGAGCTTACGTTATGTTACCTATGTTACTTTAGATTTA
S4: AAATCTAAAGAACATAAGGAACATAACGAAGCTCATTCGCG
S5: CGCGAATGAGCTTACGTTATGTTACCTTATGTTACTTTAGATTTA
The highlighted base was labeled with Cy3B (GE Healthcare) for strand S1 and S4, and with Atto647N (ATTO-TEC GmbH) for strand S2 and S3. Protected high FRET DNA standard: S1/S2, protected intermediated FRET DNA standard: S1/S3, and multi-fluorophore DNA standard: S4/S5 were chemically linked using azide–alkyne click-chemistry (Scheme 1). Experimental details and chemical structures of DNA modifications are shown in the ESI.† Protected DNA FRET standards were stored in ddH2O at −20 °C.
In vitro single-molecule FRET measurements and fluorescence correlation spectroscopy (FCS) measurements were performed on a custom-built confocal microscope.36 The confocal setup consisted of two laser lines, a 638 nm diode laser (Cube, Coherent, operated at 30 μW), and a 532 nm Nd:YAG laser (Samba, Cobolt, operated at 240 μW), which were alternated at 20 kHz for single-molecule FRET measurements and which were operated in green continuous wave-mode for FCS measurements. The laser light was coupled into a 60×, 1.35 NA, UPlanSApo oil immersion objective (Olympus) and fluorescence signal from diffusing molecules was collected by the same objective and spectrally separated by a dichroic mirror (630DRLP, Omega) and directed on two avalanche photodiodes (SPCM-AQR14, Perkin Elmer). Photon arrival times were recorded with a PC counting National Instrument board and data processing was done using custom-written LabVIEW software (National Instruments). For in vitro confocal smFRET studies 20 μL of DNA FRET standards diluted in ddH2O to 10–50 pM final concentration were added onto a burned cover slide and the laser beam was focused about 20 μm into solution; two 10 min data sets were acquired for each sample. The single-molecule fluorescence bursts from in vitro single-molecule FRET confocal microscopy were analysed in MATLAB as described in ref. 23 and as briefly described in the ESI.† For FCS measurements 20 μL of DNA FRET standards diluted in ddH2O to 1–5 nM final concentration and single-molecule bursts were recorded for 5 min per data set at 120 μW green continuous wave and separated into green and red fluorescence channel for detection. The autocorrelation analysis is described in the ESI.†
Electroporation was performed using ElectroMAX DH5α-E Competent Cells (Invitrogen). The cells were diluted 1:1 with ddH2O and stored at −80 °C. For each electroporation experiment protected DNA FRET standards were added to a final concentration of 50 nM to 20 μL of electrocompetent cells. The cell suspension was added into a pre-chilled electroporation cuvette (1 mm gap, Bio-Rad) and exposed to the discharge of a high voltage electric field with initial amplitude of 14 kV cm−1 (MicroPulser, Bio-Rad). The cells were rapidly recovered in 500 μL of super optimal broth with catabolite repression (SOC) for about 20 min shaking at 37 °C. Then, the cells were washed 4 times with phosphate buffered saline (PBS) by pelleting the cells by centrifugation at 3300g for 1 min at 4 °C, followed by resuspension. Finally, the cells were resuspended in about 100 μL PBS and stored on ice. About 5–10 μL of the cell suspension were spread on 1% agarose-M9 pads. Finally, a burned cover slide was placed on top of the agarose pad and turned towards the immersion oil objective for imaging.
In vivo single-molecule FRET measurements were performed under HILO illumination34 on a custom-built inverted widefield microscope.23 Laser light from two lasers, a 637 nm diode laser (Vortran Stradus Laser Technology) and a 532 nm DPSS laser (MGL-III-532 nm-100 mW, CNI) operated in green continuous wave-mode at 100 W cm−2 green excitation or in ALEX-mode at 100 W cm−2 green excitation, and 50 W cm−2 red excitation, was focused onto the backfocal plane of the objective and cellular fluorescence was collected through the same oil-immersion objective (UPLSAPO, 100×, NA 1.4, Olympus) and spectrally separated by a dichroic mirror (630DRLP, Omega). Each channel was imaged onto separate halves of an EMCCD camera chip (iXon+, BI-887, Andor). The illumination for brightfield images comprised a white-light lamp (IX2-ILL100, Olympus), which was attached to the microscope body. Movies and images were recorded using Andor camera software. Single-molecule FRET movies were analyzed using custom-written MATLAB software. In vivo smFRET analysis is further described in the ESI.†
Acknowledgements
We thank Timothy Craggs and Antonio Garcia Guerra for helpful discussions. A. P. was supported by a UK EPSRC DTA studentship and the German National Academic Foundation (Studienstiftung). A. P. held the Phizackerley Senior Scholarship in the Medical Sciences at Balliol College, Oxford. A. N. K. was supported by a UK BBSRC grant (BB/H01795X/1), and a European Research Council Starter grant (ERC 261227). T. B., A. N. K. and A. H. E.-S. thank the UK BBSRC for funding via the sLOLA grant BB/J001694/1: “Extending the Boundaries of Nucleic Acid Chemistry”.Notes and references
- S. J. Lord, H. L. Lee and W. E. Moerner, Anal. Chem., 2010, 82, 2192–2203 CrossRef CAS PubMed.
- X. S. Xie, P. J. Choi, G. W. Li, N. K. Lee and G. Lia, Annu. Rev. Biophys., 2008, 37, 417–444 CrossRef CAS PubMed.
- S. Weiss, Science, 1999, 283, 1676–1683 CrossRef CAS PubMed.
- T. Ha, Methods, 2001, 25, 78–86 CrossRef CAS PubMed.
- M. Sustarsic and A. N. Kapanidis, Curr. Opin. Struct. Biol., 2015, 34, 52–59 CrossRef CAS PubMed.
- T. Förster, Ann. Phys., 1948, 437, 55–75 CrossRef.
- L. Stryer and R. P. Haugland, Proc. Natl. Acad. Sci. U. S. A., 1967, 58, 719–726 CrossRef CAS.
- T. Ha, T. Enderle, D. F. Ogletree, D. S. Chemla, P. R. Selvin and S. Weiss, Proc. Natl. Acad. Sci. U. S. A., 1996, 93, 6264–6268 CrossRef CAS.
- B. Schuler, E. A. Lipman and W. A. Eaton, Nature, 2002, 419, 743–747 CrossRef CAS PubMed.
- R. Zhao and D. Rueda, Methods, 2009, 49, 112–117 CrossRef CAS PubMed.
- T. D. Christian, L. J. Romano and D. Rueda, Proc. Natl. Acad. Sci. U. S. A., 2009, 106, 21109–21114 CrossRef CAS PubMed.
- Y. Santoso, C. M. Joyce, O. Potapova, L. Le Reste, J. Hohlbein, J. P. Torella, N. D. Grindley and A. N. Kapanidis, Proc. Natl. Acad. Sci. U. S. A., 2010, 107, 715–720 CrossRef CAS PubMed.
- A. N. Kapanidis, E. Margeat, S. O. Ho, E. Kortkhonjia, S. Weiss and R. H. Ebright, Science, 2006, 314, 1144–1147 CrossRef PubMed.
- S. Arslan, R. Khafizov, C. D. Thomas, Y. R. Chemla and T. Ha, Science, 2015, 348, 344–347 CrossRef CAS PubMed.
- S. Liu, B. T. Harada, J. T. Miller, S. F. Le Grice and X. Zhuang, Nat. Struct. Mol. Biol., 2010, 17, 1453–1460 CAS.
- A. Plochowietz and A. N. Kapanidis, Nat. Methods, 2015, 12, 715–716 CrossRef CAS PubMed.
- N. C. Shaner, P. A. Steinbach and R. Y. Tsien, Nat. Methods, 2005, 2, 905–909 CrossRef CAS PubMed.
- T. Ha and P. Tinnefeld, Annu. Rev. Phys. Chem., 2012, 63, 595–617 CrossRef CAS PubMed.
- S. A. Jones, S. H. Shim, J. He and X. Zhuang, Nat. Methods, 2011, 8, 499–508 CrossRef CAS PubMed.
- A. Keppler, S. Gendreizig, T. Gronemeyer, H. Pick, H. Vogel and K. Johnsson, Nat. Biotechnol., 2003, 21, 86–89 CrossRef CAS PubMed.
- R. Wombacher, M. Heidbreder, S. van de Linde, M. P. Sheetz, M. Heilemann, V. W. Cornish and M. Sauer, Nat. Methods, 2010, 7, 717–719 CrossRef CAS PubMed.
- C. C. Liu and P. G. Schultz, Annu. Rev. Biochem., 2010, 79, 413–444 CrossRef CAS PubMed.
- R. Crawford, J. P. Torella, L. Aigrain, A. Plochowietz, K. Gryte, S. Uphoff and A. N. Kapanidis, Biophys. J., 2013, 105, 2439–2450 CrossRef CAS PubMed.
- J. J. Sakon and K. R. Weninger, Nat. Methods, 2010, 7, 203–205 CrossRef CAS PubMed.
- I. Konig, A. Zarrine-Afsar, M. Aznauryan, A. Soranno, B. Wunderlich, F. Dingfelder, J. C. Stuber, A. Pluckthun, D. Nettels and B. Schuler, Nat. Methods, 2015, 12, 773–779 CrossRef CAS PubMed.
- M. Sustarsic, A. Plochowietz, L. Aigrain, Y. Yuzenkova, N. Zenkin and A. Kapanidis, Histochem. Cell Biol., 2014, 142, 113–124 CrossRef CAS PubMed.
- A. Plochowietz, R. Crawford and A. N. Kapanidis, Phys. Chem. Chem. Phys., 2014, 16, 12688–12694 RSC.
- E. Friedberg, DNA Repair and Mutagenesis, American Society of Microbiology, 2006 Search PubMed.
- E. T. Kool, Annu. Rev. Biophys. Biomol. Struct., 1996, 25, 1–28 CrossRef CAS PubMed.
- I. K. Lee, J. D. Ahn, H. S. Kim, J. Y. Park and K. U. Lee, Curr. Drug Targets, 2003, 4, 619–623 CrossRef CAS PubMed.
- A. N. Kapanidis, N. K. Lee, T. A. Laurence, S. Doose, E. Margeat and S. Weiss, Proc. Natl. Acad. Sci. U. S. A., 2004, 101, 8936–8941 CrossRef CAS PubMed.
- N. K. Lee, A. N. Kapanidis, Y. Wang, X. Michalet, J. Mukhopadhyay, R. H. Ebright and S. Weiss, Biophys. J., 2005, 88, 2939–2953 CrossRef CAS PubMed.
- S. A. Kooijmans, S. Stremersch, K. Braeckmans, S. C. de Smedt, A. Hendrix, M. J. Wood, R. M. Schiffelers, K. Raemdonck and P. Vader, J. Controlled Release, 2013, 172, 229–238 CrossRef CAS PubMed.
- M. Tokunaga, N. Imamoto and K. Sakata-Sogawa, Nat. Methods, 2008, 5, 159–161 CrossRef CAS PubMed.
- S. J. Holden, S. Uphoff and A. N. Kapanidis, Nat. Methods, 2011, 8, 279–280 CrossRef CAS PubMed.
- J. Hohlbein, L. Aigrain, T. D. Craggs, O. Bermek, O. Potapova, P. Shoolizadeh, N. D. Grindley, C. M. Joyce and A. N. Kapanidis, Nat. Commun., 2013, 4, 2131 Search PubMed.
Footnote |
| † Electronic supplementary information (ESI) available: Experimental methods, data analysis routines and Fig. S1–S9. See DOI: 10.1039/c6sc00639f |
| This journal is © The Royal Society of Chemistry 2016 |