
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Inhibiting quinolone biosynthesis of Burkholderia†
Michaela
Prothiwa
a,
Verena
Filz
ab,
Sebastian
Oehler
 a and
Thomas
Böttcher
*ab
a and
Thomas
Böttcher
*ab
aDepartment of Chemistry, Konstanz Research School Chemical Biology, University of Konstanz, 78457 Konstanz, Germany
bFaculty of Chemistry, Department of Biological Chemistry & Centre for Microbiology and Environmental Systems Science, Division of Microbial Ecology, University of Vienna, 1090 Vienna, Austria. E-mail: Thomas.Boettcher@univie.ac.at
First published on 26th March 2021
Abstract
2-Alkylquinolones are important signalling molecules of Burkholderia species. We developed a substrate-based chemical probe against the central quinolone biosynthesis enzyme HmqD and applied it in competitive profiling experiments to discover the first known HmqD inhibitors. The most potent inhibitors quantitatively blocked quinolone production in Burkholderia cultures with single-digit micromolar efficacy.
2-Alkylquinolones (AQs) and their derivatives are important bacterial metabolites. Over 50 different AQs have been reported in Pseudomonas aeruginosa1 with major roles for virulence control via quorum sensing,2,3 interspecies competition,4,5 and bacteria–host interactions.6 Homologous biosynthetic gene clusters also exist in species of the genus Burkholderia7 producing unique methylated quinolone congeners.8,9 These 3-methyl-2-alkylquinolones (MAQs)‡ are non-classical quorum sensing signals in Burkholderia and have been implicated in regulating homoserine lactone quorum signalling.10
The genus Burkholderia comprises important pathogens such as B. pseudomallei, the causative agent of melioidosis11 and species of the B. cepacia complex including B. ambifaria and B. multivorans which are responsible for severe infections in cystic fibrosis (CF) patients.12 MAQ production was only reported in clinical strains of B. ambifaria and was absent in virulence attenuated variants as well as in environmental isolates.13 This may suggest potential roles of these quinolones in pathogenicity. Customized inhibitors of quinolone biosynthesis could thus provide valuable tools for dissecting hitherto unknown functions of quinolones in Burkholderia. We have recently described a live-cell profiling strategy with electrophilic chemical probes that led to highly effective inhibitors of quinolone biosynthesis of P. aeruginosa.14 We now take this strategy one step further to interrogate the function of HmqD, the postulated homologous quinolone synthetase of Burkholderia and discovered the first inhibitors of the quinolone production in live cells of this genus.
While the pqsA-E quinolone gene cluster of P. aeruginosa has been investigated in great detail over last years, species of the genus Burkholderia presumably use the gene products of hmqA-G for the biosynthesis of MAQs (Fig. 1a). The genes of the hmq cluster have been characterized by their homologies with genes of the pqs cluster and several of them also by analysis of knockout mutants.8 A central step in the biosynthesis of the quinolones of P. aeruginosa is the production of a coenzyme A-activated 2′-aminobenzoylacetate (2-ABA-CoA). The Claisen condensation of anthraniloyl-CoA with malonly-CoA is catalysed by the enzyme PqsD, for which we have previously established simple α-chloroacetamides with a terminal alkyne as chemical probes.15 Using competitive labelling, we could identify potent live-cell PqsD inhibitors which were based on the anthranilic acid core of the native substrate (Fig. S1†).
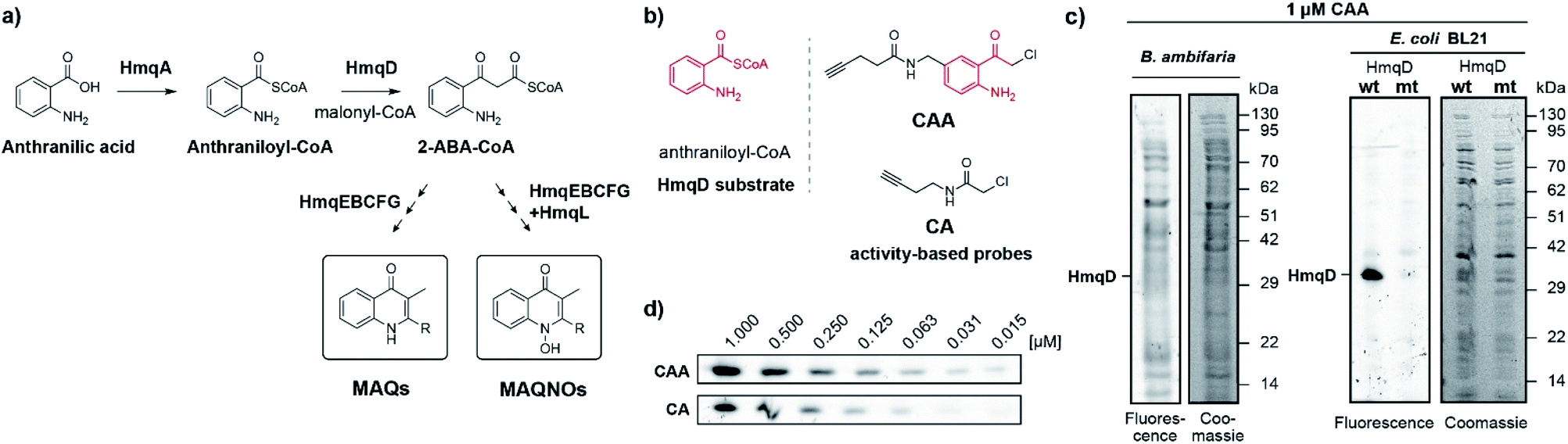 | ||
| Fig. 1 (a) Proposed pathway for the biosynthesis of quinolones by Burkholderia species, where HmqD catalyses the Claisen-type condensation reaction of anthraniloyl-CoA with malonyl-CoA to yield 2-ABA-CoA. (b) Probe design and structural analogy of probe CAA with the native HmqD substrate and comparison to the previous probe CA. (c) Fluorescent and Coomassie stained gels of proteomes of live B. ambifaria and E. coli overexpressing HmqD labelled with 1 μM of probe CAA. (d) Concentration-dependent labelling of live E. coli cells overexpressing HmqD. Wt = wild type HmqD, mt = Cys114Ala mutant HmqD. | ||
We now aimed to synthesize a more specific probe as substrate analogue for HmqD equipped with a terminal alkyne handle for attachment of a fluorescence tag by click chemistry (Fig. 1b). In short, 4-aminobenzylamine was reacted with chloroacetonitrile by Houben–Hoesch acylation to the corresponding α-chloroketone (2). Amide coupling with 4-pentynoic acid yielded probe CAA (Scheme 1).
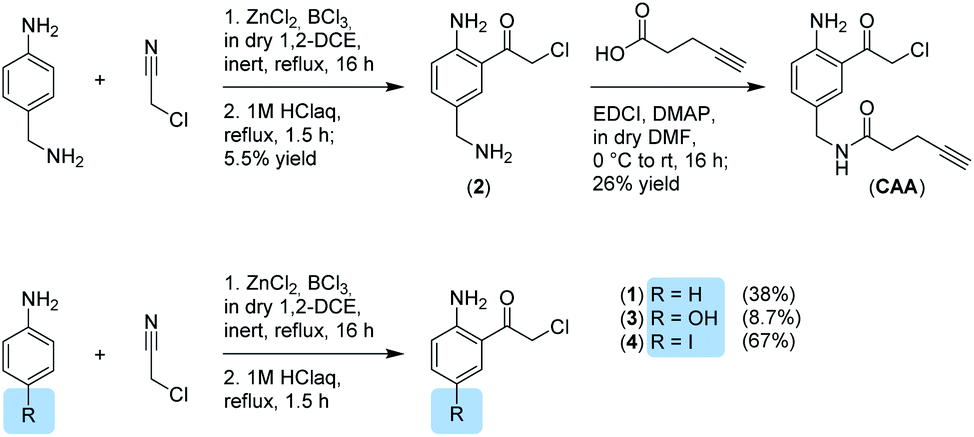 | ||
| Scheme 1 Synthesis of α-chloroketones. | ||
Live cells of Burkholdera ambifaria were incubated with probe for 30 min. Unbound probe was removed by washing steps and after cell lysis, a fluorescent tetramethylrhodamine tag was appended via bioorthogonal Cu(I) catalysed azide–alkyne cycloaddition. Subsequently, proteins were separated via sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) to allow in-gel fluorescence analysis. Expression levels of HmqD in Burkholderia ambifaria were likely too low to result in any fluorescent labelling above background (Fig. 1c).
In accordance, growth phase-resolved proteomic experiments with and without enrichment by pull-down failed to detect any peptides of HmqD suggesting very low abundance of the protein in the native proteome (ESI†). We thus generated an Escherichia coli BL21 clone recombinantly expressing HmqD of B. ambifaria as model system (Tables S1–S3†). Indeed, labelling of live cells of E. coli overexpressing HmqD with 1 μM probe CAA resulted in only one major fluorescent band. In contrast, an HmqD mutant with the active site cysteine replaced by alanine (Cys114Ala) was not labelled (Fig. 1c). This indicated not only remarkable selectivity of the probe for HmqD in the background of a live cell proteome but also excellent specificity for the active site nucleophile. Testing the CA and CAA probes side by side in concentration dependence demonstrated that labelling of HmqD in live E. coli cells was more sensitive with the new CAA probe than with the original CA probe and a fluorescent band was still detectable at only 15 nM of CAA probe (Fig. 1d and S2†). In addition, proteomic experiments with recombinant HmqD expressed in E. coli identified as site of modification by probe CAA only the active site cysteine C114 out of in total 8 cysteines of HmqD (Fig. S3†). These results validated the CAA probe as tool for competitive live cell profiling of potential HmqD inhibitors (Fig. 2a). We synthesized several α-chloroketones (1–4) using the Houben–Hoesch reaction (Scheme 1) and combined them with commercially available compounds (5–24) to generate a small library (Fig. 2b). These compounds were screened at 10 μM by pre-incubation with live cells of E. coli expressing HmqD followed by addition of 1 μM CAA probe, click chemistry and in-gel fluorescence imaging (Fig. 2a). Competitive inhibitors with sufficient cell permeability prevented the labelling by the probe resulting in reduced or complete absence of the HmqD fluorescent band in the background of the live cell proteome. Interestingly, all α-bromoketones were inactive while various α-chloroketones completely inhibited labelling at 10 μM (Fig. 2c and S4†). At higher a concentration of 50 μM, three α-bromoketones inhibited labelling of HmqD in vitro but only one showed activity in live cells, suggesting that α-bromoketones may be scavenged by off-targets binding (Fig. S5†).
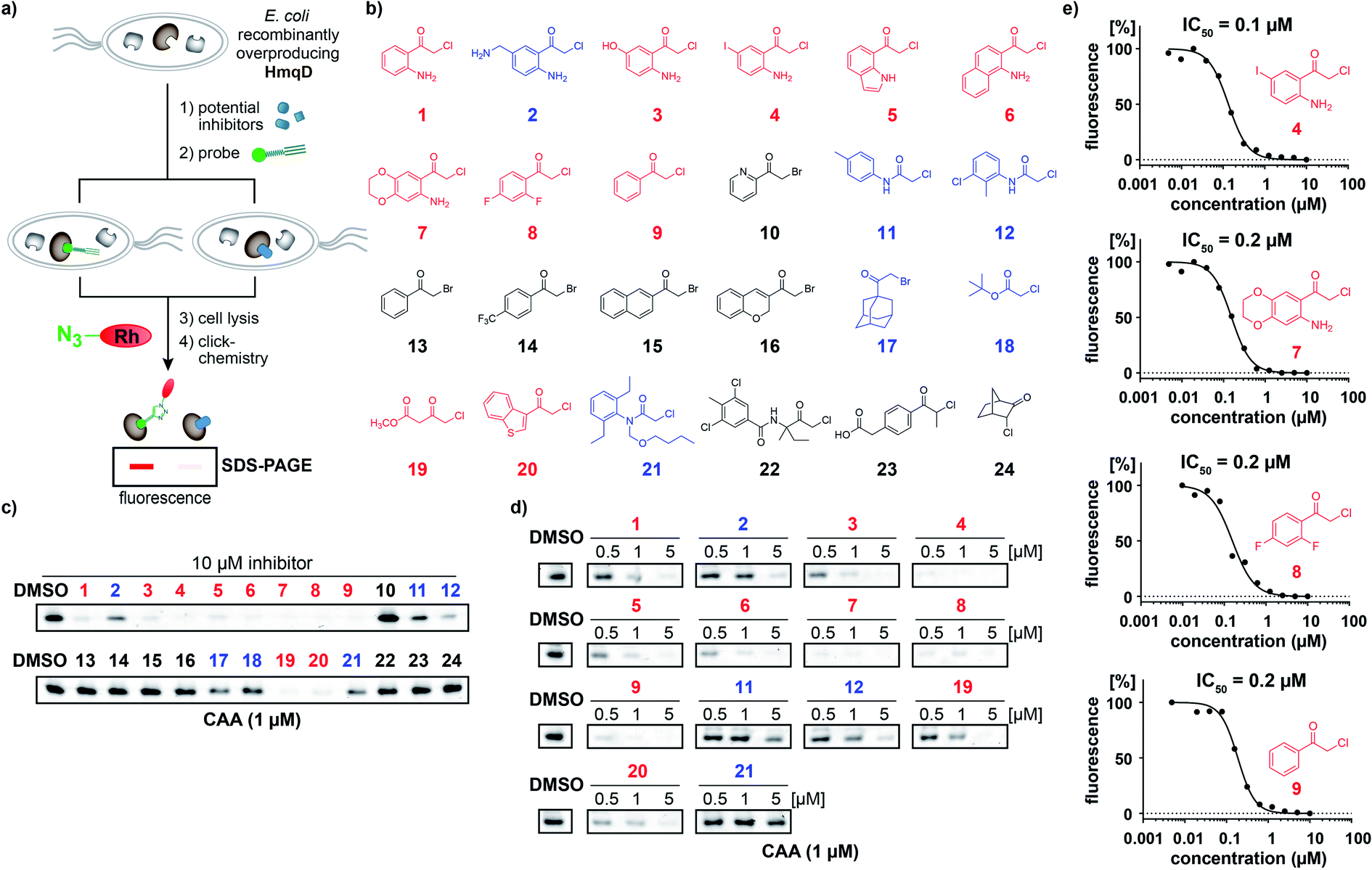 | ||
| Fig. 2 Competitive profiling for HmqD inhibitors. (a) Strategy of live-cell competitive inhibitor discovery using an activity-based probe to screen for active site-directed inhibitors of the target enzyme HmqD. (b) Structures of a focussed compound library featuring electrophilic α-chloroketone, α-chloroamide, and α-bromoketone warheads. (c) Initial competitive live-cell screening of the compound library at 10 μM against HmqD. (d) Dose-down experiments of the preliminary hits identified in (c). (e) Live-cell IC50 values for competitive HmqD inhibition. Color code: active compounds with inhibition of competitive labelling ≤1 μM (red), compounds with inhibition ≥5 μM (blue), compounds inactive at 10 μM (black). | ||
The most effective inhibitors were subjected to dose-down experiments with 5 μM, 1 μM and 0.5 μM (Fig. 2d and S6†). Four of them considerably inhibited labelling even at the lowest concentration of 0.5 μM (Fig. 2d). For these compounds a wider range of concentration was used and IC50 values were determined by quantification of labelling intensities. The anthranilic acid-derived compounds 7, 8, and 9 inhibited labelling with IC50 values of 0.2 μM and the most active 5′-iodo-substituted compound 4 even resulted in an IC50 of 0.1 μM (Fig. 2e and S7†).
The three most active inhibitors 4, 7 and 8 were finally tested in cultures of B. ambifaria strain AMMD, which has been reported to produce MAQs.8 We quantified levels of the six major MAQ derivatives in the culture supernatants using multiple reaction monitoring (MRM) on a triple quadrupole mass spectrometer measuring selective mass transitions of the most intense ions resulting from the fragmentation of precursor ions (Fig. 3a, S8, S9 and Table S4†).8 Isolated and purified Δ2MHQ was spiked into the growth medium and served as quantification standard to confirm linearity within the detection range (Table S4†). Strikingly, after growing cultures of B. ambifaria in presence of inhibitors for 24 h under oxygen limiting conditions, compound 4 and 7 completely blocked MAQ production at concentrations as low as 5 μM (Fig. 3b, S10 and S11†). In contrast, compound 8 only abolished global MAQ production in a higher concentration of 50 μM. Growth was not affected by the most active inhibitor 4 at a concentration of 50 μM over an incubation period of 40 h (Fig. 3c), confirming that low MAQ levels are not artefacts of growth inhibition. Interestingly, under aerobic conditions, MAQ concentrations of B. ambifaria peaked in the late exponential growth phase within a narrow time frame of 3 h (Fig. 3c and S12†). The sharp peak and the subsequent complete disappearance of MAQs in culture supernatants indicate a tightly regulated production and suggest the existence of a degradation mechanism. Under oxygen limiting conditions, MAQ levels were considerably lower and continuously produced over a longer period of time in the late stationary growth phase, which implies a modulation of the MAQ biosynthesis by oxygen.
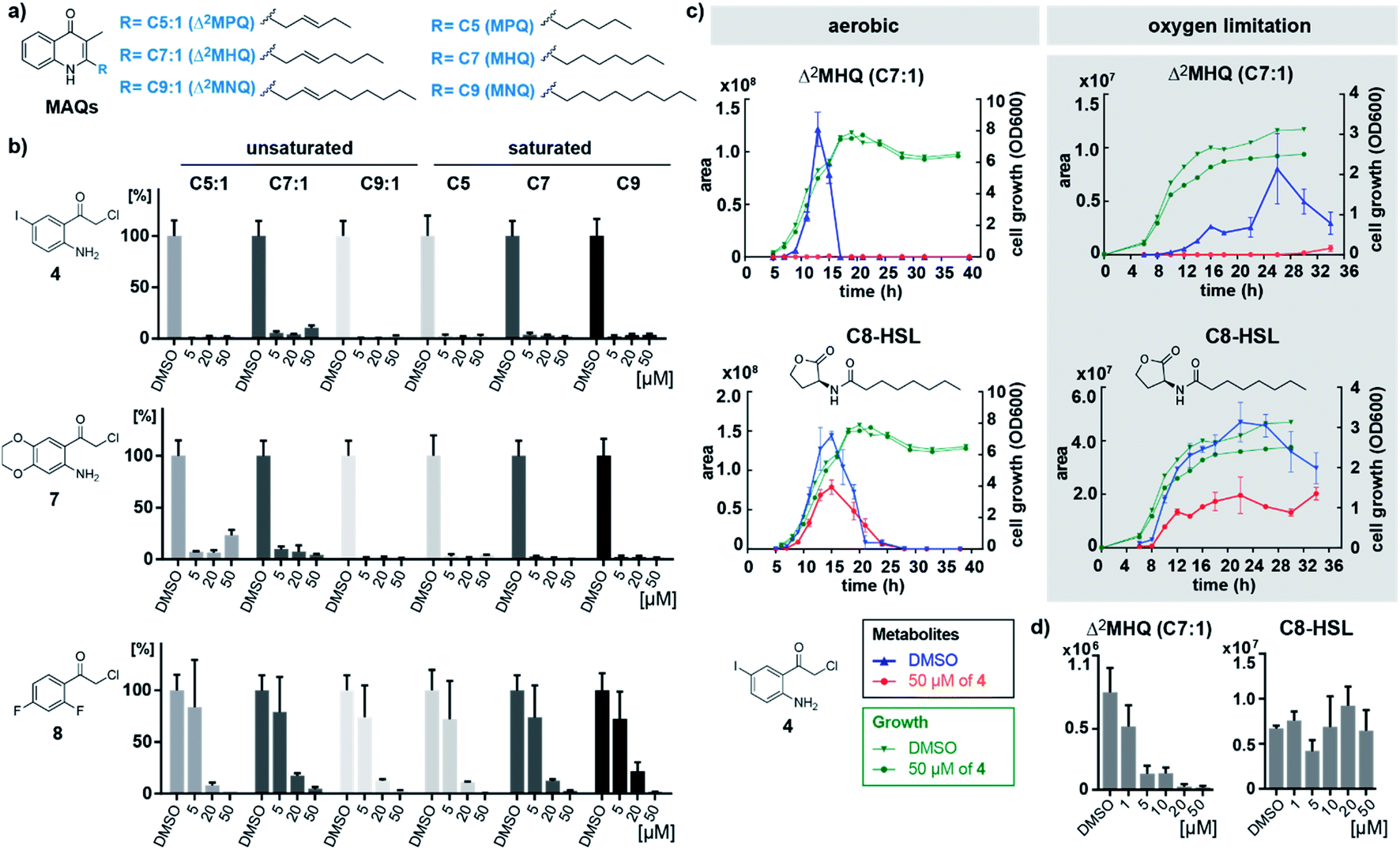 | ||
| Fig. 3 (a) Structures of unsaturated and saturated 3-methyl-2-alkylquinolones (MAQs) with different chain lengths naturally produced by B. ambifaria. (b) MAQs detected in the culture supernatants of B. ambifaria treated for 24 h with different concentrations of inhibitors 4, 7 and 8 under oxygen limiting conditions. Bars represent normalized integrated areas of recorded mass transitions with standard deviation of experiments performed in biological triplicates. (c) Overlay of growth curves and time-resolved mass spectrometric detection of the main quinolone Δ2MHQ and the N-octanoyl-L-homoserine lactone (C8-HSL) quorum sensing signal of B. ambifaria grown under aerobic and oxygen limiting culture conditions with 50 μM of inhibitor 4 in comparison to DMSO control. (d) Concentration-dependence of the effect of inhibitor 4 on Δ2MHQ and C8-HSL detected in B. ambifaria cultures after 11 h under aerobic conditions. | ||
To investigate the effect of inhibitors on other quorum sensing systems, we quantified acyl homoserine lactones (AHLs) in the supernatant of B. ambifaria AMMD (Table S5†).
Under aerobic conditions highest AHL concentrations were detected between 10 and 20 h of growth, with 3-OH-C10-HSL and C8-HSL as mainly produced AHLs (Fig. S13†). Oxygen limiting conditions resulted in significantly lower AHL concentrations with maximal production in the stationary phase. It is remarkable that production levels as well as timing of quinolone and AHL production appear to be strictly controlled by oxygen levels. Both conditions (normoxic and anoxic) are encountered in the lungs of cystic fibrosis patients.16 Consistent with our findings, species of the Burkholderia cepacia complex are known to adaptively respond to anaerobic conditions in the cystic fibrosis lung.17 While MAQ production was completely inhibited by compound 4, AHL production was only mildly affected. To confirm this specificity, we profiled MAQ and AHL levels in culture supernatants of B. ambifaria grown for 11 h under aerobic conditions treated with a range of concentrations of compound 4. MAQ production was considerably inhibited already at 1 μM of 4 and further decreased with higher concentrations. In contrast, AHLs showed no concentration-dependent inhibition (Fig. 3d and S15†). These results demonstrate that compound 4 is specific for inhibition of quinolone biosynthesis in B. ambifaria AMMD.
To detect potential off-targets of inhibitor 4, we conducted additional competitive labelling experiments. Proteomic experiments revealed several potential off-targets of the CAA probe (Tables S6 and S7†) and we additionally generated a reactive α-bromoacetamide probe (BA). Even at 10-fold excess, compound 4 did not compete noticeably with the off-target labelling by probe CAA or BA (Fig. S14†), providing further support for a certain level of specificity of inhibitor 4.
Our results not only show the value of competitive profiling of live cells with activity-based probes, but to the best of our knowledge also provide the first inhibitors of quinolone biosynthesis in the genus Burkholderia. Inhibiting quinolone signalling of Pseudomonas aeruginosa is an important anti-infective strategy aiming to ameliorate virulence and pathogenicity.14,18,19 While in P. aeruginosa 2-alkylquinolones (HHQ and PQS) are important quorum sensing signals,20,21 recent work suggested that MAQs do not induce their own production in B. ambifaria strain HSJ1 and are probably acting on and regulated via the CepI/CepR AHL quorum sensing system.10 Although the exact roles of MAQs in the Burkholderia cepacia complex still need to be investigated, the homologous AQs of P. aeruginosa indicate possible roles in suppressing the host immune response, mediating the killing of host cells and modulating interspecies and interkingdom interactions.22–24
Conclusions
In conclusion, we show that a substrate-analogous activity-based probe targeting the central quinolone biosynthesis enzyme HmqD of B. ambifaria can be applied to find competitive inhibitors in a live cell model and present the first known HmqD inhibitors. The most potent HmqD inhibitors are able to completely block quinolone biosynthesis in cultures of B. ambifaria even in single-digit micromolar concentrations, while keeping the production of homoserine lactone quorum sensing signals intact. Thus, our compounds represent promising chemical tools for dissecting the role of quinolones in pathogenicity of Burkholderia species.Author contributions
M. P. and T. B. conceived and designed the study. M. P., V. F., and S. O. synthesized the probes, produced the recombinant proteins, and performed the labelling experiments. M. P. and V. F. extracted and analyzed the metabolites by mass spectrometry. M. P., V. F., S. O., and T. B. analyzed and interpreted the data. M. P. and T. B. wrote the manuscript.Conflicts of interest
There are no conflicts to declare.Acknowledgements
We thank Prof. Dr Andreas Marx for his generous support and Malin Bein and Prof. Dr Tanja Gaich for access to mass spectrometry. We gratefully acknowledge funding by the Emmy Noether program of the Deutsche Forschungsgemeinschaft (DFG), EU FP7 Marie Curie Zukunftskolleg Incoming Fellowship Program – University of Konstanz grant no. 291784, SFB969 project C06, and Konstanz Research School Chemical Biology (KoRS-CB). MP was supported by a Zeiss PhD fellowship.Notes and references
- F. Lepine, S. Milot, E. Deziel, J. He and L. G. Rahme, J. Am. Soc. Mass Spectrom., 2004, 15, 862–869 CrossRef CAS PubMed.
- S. McGrath, D. S. Wade and E. C. Pesci, FEMS Microbiol. Lett., 2004, 230, 27–34 CrossRef CAS.
- D. Maura, R. Hazan, T. Kitao, A. E. Ballok and L. G. Rahme, Sci. Rep., 2016, 6, 34083 CrossRef CAS PubMed.
- D. Szamosvári and T. Böttcher, Angew. Chem., 2017, 56, 7271–7275 CrossRef PubMed.
- D. Szamosvári and T. Böttcher, Synlett, 2018, 29, 542–547 CrossRef.
- D. S. Hooi, B. W. Bycroft, S. R. Chhabra, P. Williams and D. I. Pritchard, Infect. Immun., 2004, 72, 6463–6470 CrossRef CAS PubMed.
- S. P. Diggle, P. Lumjiaktase, F. Dipilato, K. Winzer, M. Kunakorn, D. A. Barrett, S. R. Chhabra, M. Camara and P. Williams, Chem. Biol., 2006, 13, 701–710 CrossRef CAS PubMed.
- L. Vial, F. Lepine, S. Milot, M. C. Groleau, V. Dekimpe, D. E. Woods and E. Deziel, J. Bacteriol., 2008, 190, 5339–5352 CrossRef CAS PubMed.
- D. Szamosvári, M. Prothiwa, C. L. Dieterich and T. Böttcher, Chem. Commun., 2020, 56, 6328–6331 RSC.
- A. Chapalain, M. C. Groleau, S. Le Guillouzer, A. Miomandre, L. Vial, S. Milot and E. Deziel, Front. Microbiol., 2017, 8, 1021 CrossRef PubMed.
- W. J. Wiersinga, H. S. Virk, A. G. Torres, B. J. Currie, S. J. Peacock, D. A. B. Dance and D. Limmathurotsakul, Nat. Rev. Dis. Primers, 2018, 4, 17107 CrossRef PubMed.
- V. C. Scoffone, L. R. Chiarelli, G. Trespidi, M. Mentasti, G. Riccardi and S. Buroni, Front. Microbiol., 2017, 8, 1592 CrossRef PubMed.
- L. Vial, M. C. Groleau, M. G. Lamarche, G. Filion, J. Castonguay-Vanier, V. Dekimpe, F. Daigle, S. J. Charette and E. Deziel, ISME J., 2010, 4, 49–60 CrossRef PubMed.
- M. Prothiwa, F. Englmaier and T. Böttcher, J. Am. Chem. Soc., 2018, 140, 14019–14023 CrossRef CAS PubMed.
- M. Prothiwa, D. Szamosvari, S. Glasmacher and T. Böttcher, Beilstein J. Org. Chem., 2016, 12, 2784–2792 CrossRef CAS PubMed.
- E. S. Cowley, S. H. Kopf, A. LaRiviere, W. Ziebis and D. K. Newman, mBio, 2015, 6, e00767 CrossRef CAS PubMed.
- U. Schwab, L. H. Abdullah, O. S. Perlmutt, D. Albert, C. W. Davis, R. R. Arnold, J. R. Yankaskas, P. Gilligan, H. Neubauer, S. H. Randell and R. C. Boucher, Infect. Immun., 2014, 82, 4729–4745 CrossRef PubMed.
- C. Schütz and M. Empting, Beilstein J. Org. Chem., 2018, 14, 2627–2645 CrossRef PubMed.
- M. P. Storz, C. K. Maurer, C. Zimmer, N. Wagner, C. Brengel, J. C. de Jong, S. Lucas, M. Musken, S. Haussler, A. Steinbach and R. W. Hartmann, J. Am. Chem. Soc., 2012, 134, 16143–16146 CrossRef CAS PubMed.
- S. P. Diggle, S. Matthijs, V. J. Wright, M. P. Fletcher, S. R. Chhabra, I. L. Lamont, X. Kong, R. C. Hider, P. Cornelis, M. Camara and P. Williams, Chem. Biol., 2007, 14, 87–96 CrossRef CAS PubMed.
- H. Huse and M. Whiteley, Chem. Rev., 2011, 111, 152–159 CrossRef CAS PubMed.
- K. Kim, Y. U. Kim, B. H. Koh, S. S. Hwang, S. H. Kim, F. Lepine, Y. H. Cho and G. R. Lee, Immunology, 2010, 129, 578–588 CrossRef CAS PubMed.
- F. J. Reen, M. J. Mooij, L. J. Holcombe, C. M. McSweeney, G. P. McGlacken, J. P. Morrissey and F. O'Gara, FEMS Microbiol. Ecol., 2011, 77, 413–428 CrossRef CAS PubMed.
- G. D. Vrla, M. Esposito, C. Zhang, Y. Kang, M. R. Seyedsayamdost and Z. Gitai, PLoS Pathog., 2020, 16, e1008867 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d0sc06167k |
| ‡ 3-Methyl-2-alkylquinolones (MAQs) have been also termed HMAQs referring to their tautomeric 4-hydroxy-3-methyl-2-alkyl-4(1H)-quinolone form. To be consistent with the nomenclature of 2-alkyl-4(1H)-quinolones (AQs), we chose the abbreviation MAQs and MAQNOs for the corresponding N-oxides. |
| This journal is © The Royal Society of Chemistry 2021 |