
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceContributions of methionine to recognition of trimethyllysine in aromatic cage of PHD domains: implications of polarizability, hydrophobicity, and charge on binding†
Katherine I.
Albanese
 and
Marcey L.
Waters
*
and
Marcey L.
Waters
*
Department of Chemistry, University of North Carolina at Chapel Hill, CB 3290, Chapel Hill, NC 27599, USA. E-mail: mlwaters@email.unc.edu
First published on 2nd June 2021
Abstract
Recognition of trimethyllysine (Kme3) by reader proteins is an important regulator of gene expression. This recognition event is mediated by an aromatic cage made up of 2–4 aromatic residues in the reader proteins that bind Kme3 via cation–π interactions. A small subset of reader proteins contain a methionine (Met) residue in place of an aromatic sidechain in the binding pocket. The unique role of sulfur in molecular recognition has been demonstrated in a number of noncovalent interactions recently, including interactions of thiols, thioethers, and sulfoxides with aromatic rings. However, the interaction of a thioether with an ammonium ion has not previously been investigated and the role of Met in binding Kme3 has not yet been explored. Herein, we systematically vary the Met in two reader proteins, DIDO1 and TAF3, and the ligand, Kme3 or its neutral analog tert-butyl norleucine (tBuNle), to determine the role of Met in the recognition of the cationic Kme3. Our studies demonstrate that Met contributes to binding via dispersion forces, with about an equal contribution to binding Kme3 and tBuNle, indicating that electrostatic interactions do not play a role. During the course of these studies, we also discovered that DIDO1 exhibits equivalent binding to tBuNle and Kme3 through a change in the mechanism of binding.
Introduction
Trimethyllysine (Kme3) is a post-translational modification (PTM) that regulates chromatin remodeling and gene expression.1–4 It functions by creating a new binding epitope for reader proteins that further recruit protein complexes to the nucleosome to elicit a biological response.5–7 Over 170 methyllysine reader proteins are known in humans, and development of inhibitors and molecular probes for these proteins is an active area of research,8 as dysregulation of lysine methylation has been associated with a wide range of diseases.3,9 An aromatic cage made up of 2–4 aromatic residues is the ubiquitous binding motif in these Kme3 reader domains.4,10–17 Many studies have validated the necessity of the aromatic side chains for providing specific Kme interactions through mutagenesis efforts.4,10–17 The positively charged PTM binds to the aromatic cage via cation–π interactions, the favorable electrostatic interaction between a positive charge and the electron rich face on an aromatic ring.18,52,56 Interestingly, PHD domains diverge from other Kme3 readers in that they have a conserved methionine (Met, M) residue that occupies the back face of the cage in place of what would normally be another aromatic residue (Fig. 1).13,19–27 In contrast, there are no examples of a reader protein with an aliphatic residue (Ala, Val, Leu, or Ile) in the aromatic cage, suggesting a unique role for Met in binding Kme3.11,13,28 A range of noncovalent interactions with sulfur have been characterized, most notably sulfur–π interactions.29–41 Additionally, a recent study reported a Arg–Met–π triad, suggesting that cation–sulfur interactions may be favorable.29 Thus, we were interested in determining whether and how the methionine in PHD domains contributes energetically to binding Kme3 through favorable interactions with the cation or if its role is purely structural.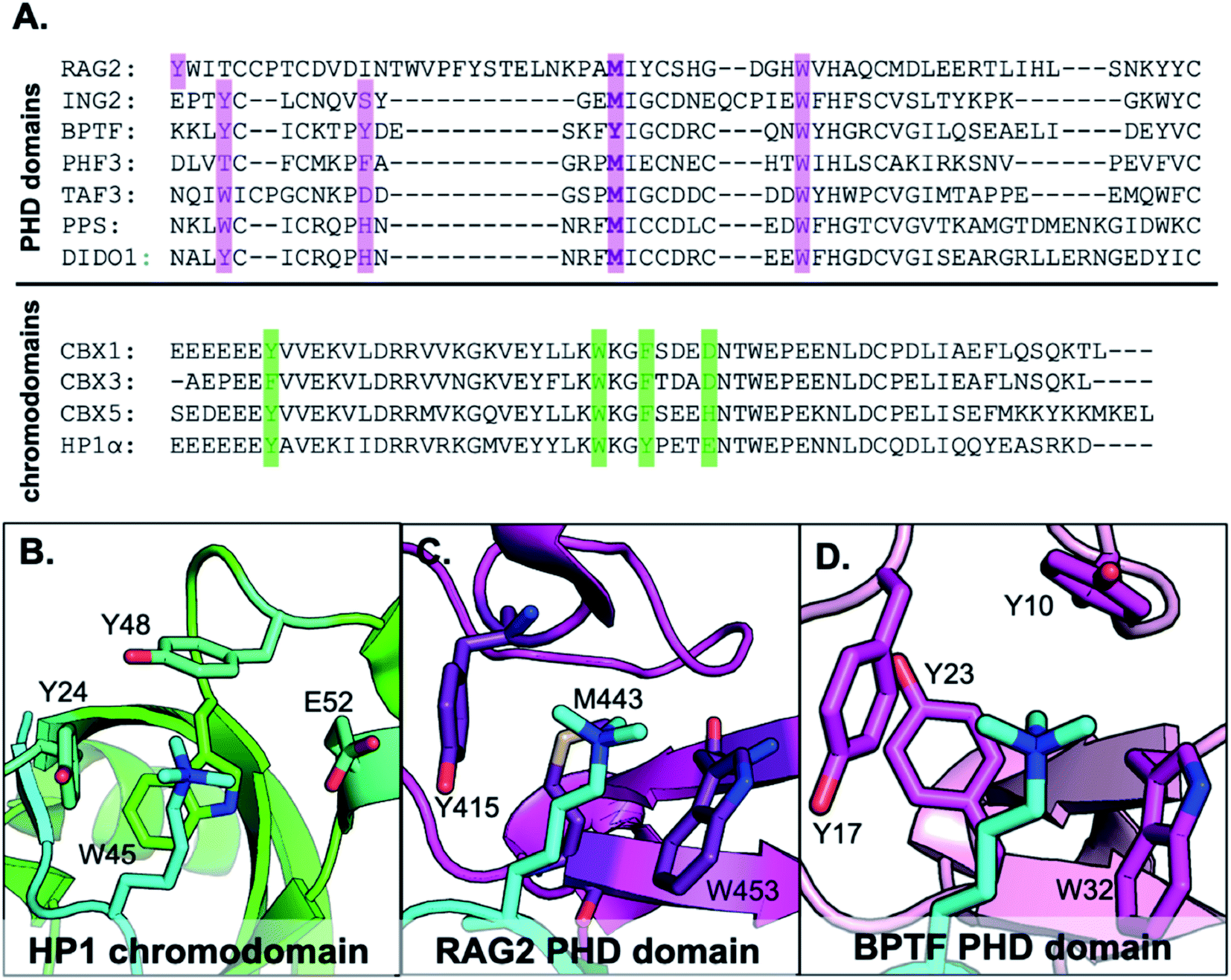 | ||
| Fig. 1 (A) Sequence alignment of PHD domains that recognize H3K4me3 and chromodomains that recognize H3K9me3. Highlighted in green are residues that make up the aromatic cage motif of chromodomains, and PHD domains in purple. (B–D) Examples of reader domain types that bind Kme3. (B) HP1 chromodomain bound to H3K9me3 with 3 aromatic residues in the binding cage (PDB ID: 1KNE). (C) RAG2 PHD domain bound to H3K4me3 with two aromatic residues and one Met in the aromatic cage (PDB ID: 2V89). (D) BPTF PHD domain (PDB ID: 2F6J). | ||
Herein, we report a mechanistic study to evaluate the role of methionine in recognition of trimethyllysine by systematically varying both the Met residue in the aromatic cage and the charge on the histone PTM. In addition to mutating Met to the canonical amino acids alanine (Ala, A), cysteine (Cys, C), and phenylalanine (Phe, F) (Table S1†), we harnessed the promiscuity of the methionine tRNA synthetase42–47 to incorporate Met isosteres norleucine (Nle) and selenomethionine (SeMet) (Fig. 2A) in two representative PHD domains, TAF3 and DIDO1 (Fig. 3). We also mutated the analogous tyrosine (Tyr, Y), Y23, in the PHD domain BPTF, an outlier in this family of reader proteins, to a methionine to compare binding to a 4-residue aromatic cage (Fig. 1D). To explore the role of the ammonium cation to the interaction with Met, Kme3 was mutated to a neutral analog, tert-butyl norleucine (tBuNle) (Fig. 2B). We found that Met contributes favorably to binding through dispersion forces rather than only playing a structural role, and that the interaction of Met with the positive charge of the Kme3 contributes minimally to binding. Finally, we found that despite the shared dispersive interactions between Met and Kme3 in these closely related PHD domains, TAF3 and DIDO1 exhibit different selectivity for Kme3 versus tBuNle, demonstrating the complexity of combinations of interactions that contribute to binding in these systems.
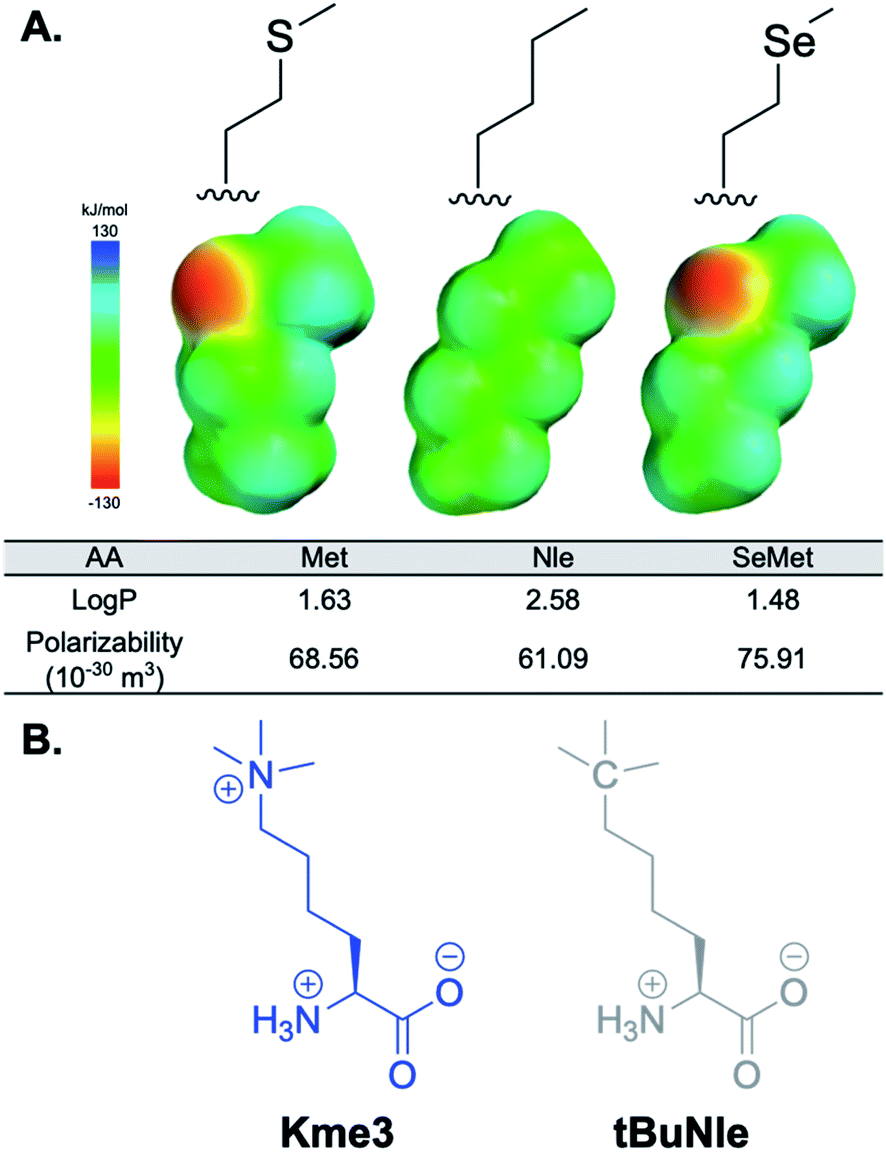 | ||
Fig. 2 (A) Analogs of methionine studied. log![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) P and polarizability values were calculated at the m06-2x/6-311+g(d,p) level of theory. (B) Chemical structures of Kme3 and tBuNle side chains. P and polarizability values were calculated at the m06-2x/6-311+g(d,p) level of theory. (B) Chemical structures of Kme3 and tBuNle side chains. | ||
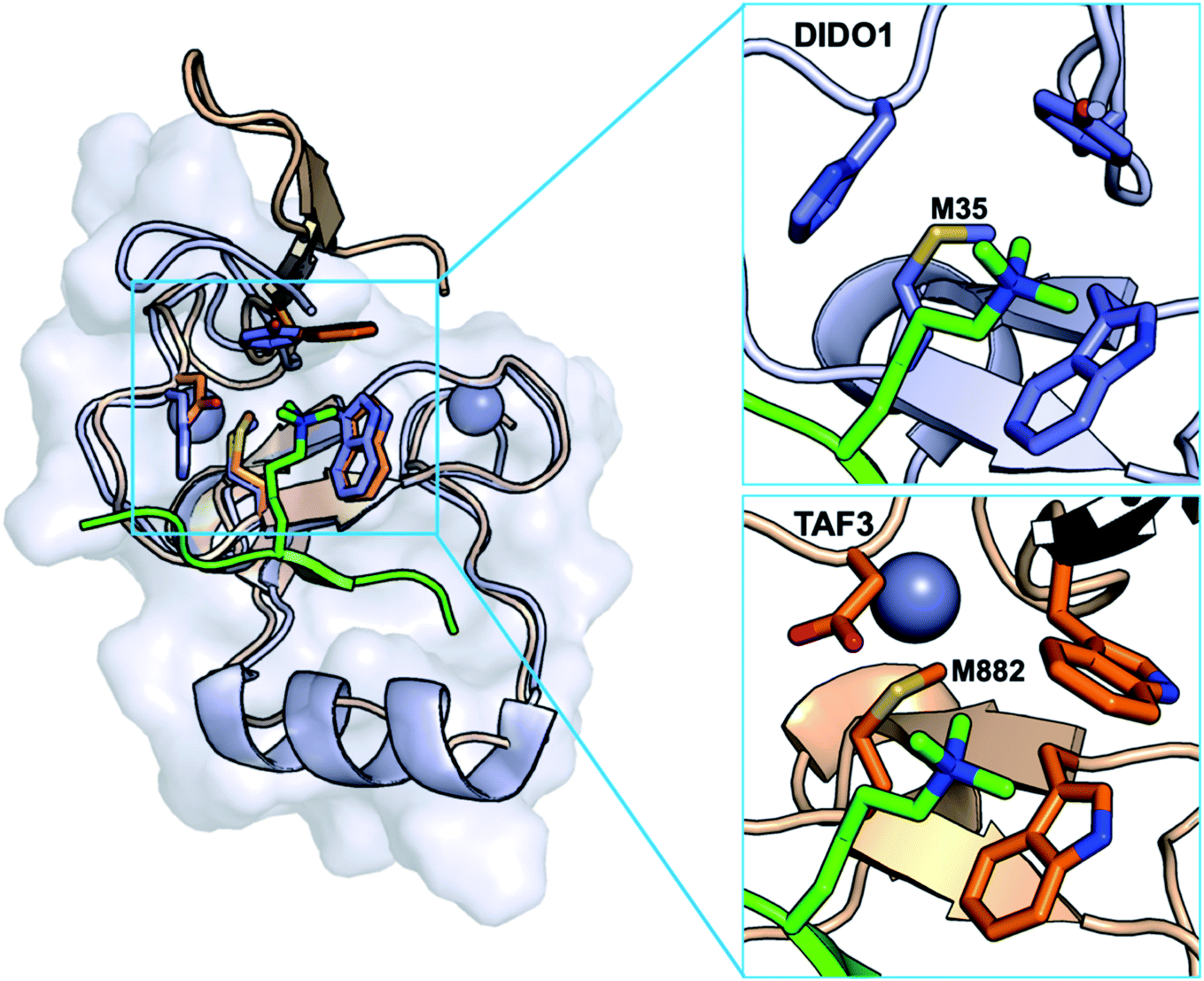 | ||
| Fig. 3 Structural alignment of DIDO1 (purple, PDB ID: 4L7X) and TAF3 (orange, PDB ID: 5WXH). Both PHD domains recognize H3K4me3 (green) through formation of a three-stranded β-sheet and cation–π interactions. Both domains contain a conserved methionine that occupies the back face of the aromatic cage. | ||
Results and discussion
Mutational strategies in model PHD domains DIDO1 and TAF3
We selected DIDO1 (death inducer obliterator 1) and TAF3 (tatabox protein associating factor 3) PHD domains, which bind histone 3 K4me3 (H3K4me3), as our model reader proteins because they both contain Met in their Kme3 binding pockets and their bound wild-type structures have been determined, which provides a starting point for considering the role of Met in binding Kme (Fig. 3). Additionally, both contain only 1–2 additional methionine residues in their primary sequences (Fig. 1A).21,25–27 This was an important determinant because incorporating Met analogs cannot be done sequence selectively as in the case of genetic code expansion technology, and we did not want confounding effects influencing the interpretation of our binding studies. To this end, we mutated M900 and M907 in TAF3 to glutamine (Gln, Q). Although both mutations slightly weakened binding, circular dichroism (CD) spectra indicated that the mutations did not affect global structure (Fig. S12†). For the purposes of this report, the TAF3 M900Q/M907Q mutant is considered the wild-type construct.Extensive work has been reported to contextualize the implications of both of these PHD domains in epigenetic regulation.21,24,27,48,49 Mutational studies of the aromatic cage by either alanine scanning or point mutation to different aromatic residues (i.e. Trp-to-Tyr/Phe) confirm the importance of cation–π interactions for selective recognition of trimethyllysine.24,27 Additional investigations into the binding sites of both proteins concluded that the histidine in DIDO1 is necessary for pH dependent recognition of H3K4me3,26 and the aspartic acid in the equivalent position in TAF3 forms important hydrogen bonds with the H3T6 side chain and a water molecule near the cage.27,50–52 In both systems, there are significant NMR chemical shift perturbations of Met upon binding H3K4me3,24,27 suggesting Met makes key contacts with the PTM. Mutation of Met to Ala resulted in loss of binding, however no additional mutational analysis has been conducted to further investigate its role.27,53
To characterize the role of Met in the aromatic cage, we investigated several canonical and noncanonical mutations of the Met within the binding pockets of DIDO1 and TAF3. Mutation to alanine was made in DIDO1 (M35A) and TAF3 (M882A) to validate previously published results demonstrating the necessity of a sidechain at the back face of the aromatic cage.24,27 Met-to-Cys has not previously been reported in either domain and was included to examine the importance of the thioether moiety. Cys is a shorter and smaller volume sidechain, but still places a sulfur in the binding pocket, and is thus a more conservative mutation than Ala.
We also investigated mutating Met-to-Phe in DIDO1 and TAF3 as it creates a complete aromatic cage in both proteins, analogous to the BPTF PHD domain, which is structurally similar (RMSD = 0.33, Fig. S15†), but has an additional aromatic residue in place of the otherwise conserved Met in the aromatic cage (Fig. 1D). Furthermore, we also mutated the tyrosine in BPTF to methionine to compare the contribution of Met vs. Tyr to binding. Of note, for each of these mutations, the optimal Φ–ψ angles in an antiparallel β-sheet match those found at the Met position in both TAF3 and DIDO1, so the conformational preferences are well aligned and should not be directly implicated in any potential unfavorable binding.54
Inspired by work from the Honek lab,42–46 Tirrell lab,47 and our own previous investigations into cation–π interactions,55 we also incorporated isosteres of Met that are structurally more analogous to Met, but have different chemical properties (Fig. 2). Norleucine (Nle) and selenomethionine (SeMet) were included to probe the effect hydrophobic interactions and polarizability have on binding. Norleucine is the most hydrophobic of the side chains investigated, which could provide favorable contacts with the N-methyls and rest of the alkyl chain of Kme3 via the hydrophobic effect. Selenium is larger, more polarizable, and less hydrophobic than sulfur, and therefore allows us to measure the influence of dispersion forces on binding to Kme3. Investigating the mutations in this systematic way provides insight into the contribution each chemical parameter has on binding.
Finally, to investigate the importance of the positive charge of Kme3, we synthesized a neutral analog, tBuNle, in place of H3K4me3 in the histone tail peptide for binding studies. This unnatural amino acid has been previously used in our lab and others to probe the necessity of the positive charge on reader domain–histone tail recognition to demonstrate that the cation–π interaction was essential for binding.52,56 The positive charge notwithstanding, both Kme3 and tBuNle are equally polarizable; if dispersion forces dominate the Met–Kme3 interaction, then tuning the interaction should have an equal effect on binding to tBuNle. The histone tail peptide consisted of amino acid residues 1–12 of H3 (ARTXQTARKSTGGY, X = Kme3 or tBuNle) as they contain both the modification site of interest (K4) as well as the sequence selective recognition motif (residues 1–6) and a GGY sequence at the C-terminus for concentration determination.
Canonical mutations to DIDO1 and TAF3 show variable effects on binding to H3K4me3
Binding of the H3K4me3 peptide to DIDO1 and TAF3, as measured by ITC, was found to be in good agreement with previously reported binding affinities (Table 1) for both wild-type constructs and the TAF3 M882A mutant.24,27,48 Mutation of methionine to alanine was unstable in DIDO1, as the protein was never successfully purified, consistent with prior studies.24 Though TAF3 M882A has been demonstrated to be a loss of function mutation through histone protein pulldown experiments,21 its affinity for H3K4me3 has not previously been reported. We determined binding of the TAF3 M882A mutant to be >100-fold weaker, although it did not appear to perturb the overall global structure based on CD (Table 1 and Fig. S12†).| Mutationa | K d (μM) | ΔG° (kcal mol−1) | ||
|---|---|---|---|---|
| DIDO1 | TAF3 | DIDO1 | TAF3 | |
| a Experiments were performed by titrating H3K4me3 peptide (500 μM to 3 mM) into TAF3 and DIDO1 mutants (50–200 μM) in 50 mM sodium phosphate, pH 7.4, 150 mM NaCl, 2 mM TCEP at 25 °C. Error was determined as standard error of the mean. b N = 1. | ||||
| Met | 2.28 ± 0.03 | 1.05 ± 0.01 | −7.70 ± 0.01 | −8.16 ± 0.01 |
| Ala | nd | 120 ± 50 | nd | −5.4 ± 0.3 |
| Cys | 36 ± 1 | nd | −6.07 ± 0.01 | nd |
| Pheb | nd | >120 | nd | <−5.4 |
The cysteine mutation was stable in DIDO1 but was not in TAF3 as the protein was not successfully isolated (Table 1). DIDO1 M35C exhibits ∼15-fold weaker binding to H3K4me3, suggesting that the thioether in Met contributes to binding. Adding to the nuanced differences between these two domains, mutating Met to Phe to closely mimic BPTF resulted in no expressed full-length protein in the case of DIDO1, while expression and binding was successfully characterized for the TAF3 M882F mutant (Table 1). This mutation disrupted binding, possibly by Phe taking on a conformation that fills the binding site which then obstructs Kme3 from interacting. These results suggest that Met has a structural role and contributes directly to binding, such that even when well folded, mutation at this position alters the binding affinity.
Given that BPTF did not evolve to contain a methionine in its aromatic cage, we explored how mutation of Tyr-to-Met (Y23M) would affect binding to H3K4me3. Interestingly, mutation in BPTF to Met was stable, and binding was only 3-fold weaker for Kme3 than what is reported for wild-type protein (7.5 μM vs. 2.7 μM,57 respectively, Table S6†) amounting only about 0.65 kcal mol−1 loss in binding. This relatively small loss in affinity suggests that Met may partially compensate for the loss of the cation–π interaction with Tyr through favorable interactions with Kme3.
Evaluation of driving forces for interaction of Met with Kme3
From the results of the Ala, Cys, and Phe mutations in DIDO1 and TAF3, it is clear that Met contributes to recognition of H3K4me3. However, these canonical mutations alone do not elucidate the role the thioether may be playing in direct interaction with Kme3 in DIDO1 and TAF3. Met is often considered to be a hydrophobic sidechain, providing a structural role within a protein core, but as has been shown in other studies, its unique properties clearly contribute more than just the hydrophobic effect.58 Sulfur is very polarizable and its participation in noncovalent interactions are predominantly dispersive in nature and can be electrostatically favorable as well.29–31 Nle, the carbon analog of Met, is more hydrophobic and less polarizable and dispersive (Fig. 2). If Met contributes to binding of Kme3 via the hydrophobic effect in PHD domains, one would expect binding to Nle mutants to be more favorable. SeMet is more polarizable than Met due to its larger size but is otherwise similar (Fig. 2). If dispersion forces are key contributors to the Met–Kme3 interaction, then binding to the SeMet mutants should be more favorable.To characterize more specifically the chemical nature of the Met–Kme3 interaction, Nle and SeMet mutations were successfully expressed and purified in both domains, and binding affinities were measured using ITC (Table 2). Comparison of the Met, Nle, and SeMet affinities in DIDO1 and TAF3 indicate that DIDO1 is more sensitive to the mutations than TAF3. Moreover, TAF3 exhibits a trend with Nle < Met < SeMet, whereas for DIDO1, Met has tighter binding to Kme3 than Nle and SeMet.
| Mutationa | K d (μM) | ΔG° (kcal mol−1) | ΔΔG° (kcal mol−1) | |||
|---|---|---|---|---|---|---|
| DIDO1 | TAF3 | DIDO1 | TAF3 | DIDO1 | TAF3 | |
| a Experiments were performed by titrating H3K4me3 peptide (500 μM to 1 mM) into TAF3 and DIDO1 mutants (50–200 μM) in 50 mM sodium phosphate, pH 7.4, 150 mM NaCl, 2 mM TCEP at 25 °C. Error was determined as standard error of the mean. | ||||||
| Met | 2.28 ± 0.03 | 1.05 ± 0.01 | −7.70 ± 0.01 | −8.16 ± 0.01 | — | — |
| Nle | 10 ± 2 | 1.7 ± 0.2 | −6.9 ± 0.1 | −7.90 ± 0.06 | 0.9 ± 0.2 | 0.3 ± 0.2 |
| SeMet | 4.2 ± 0.2 | 0.84 ± 0.07 | −7.34 ± 0.02 | −8.29 ± 0.05 | 0.36 ± 0.05 | −0.19 ± 0.09 |
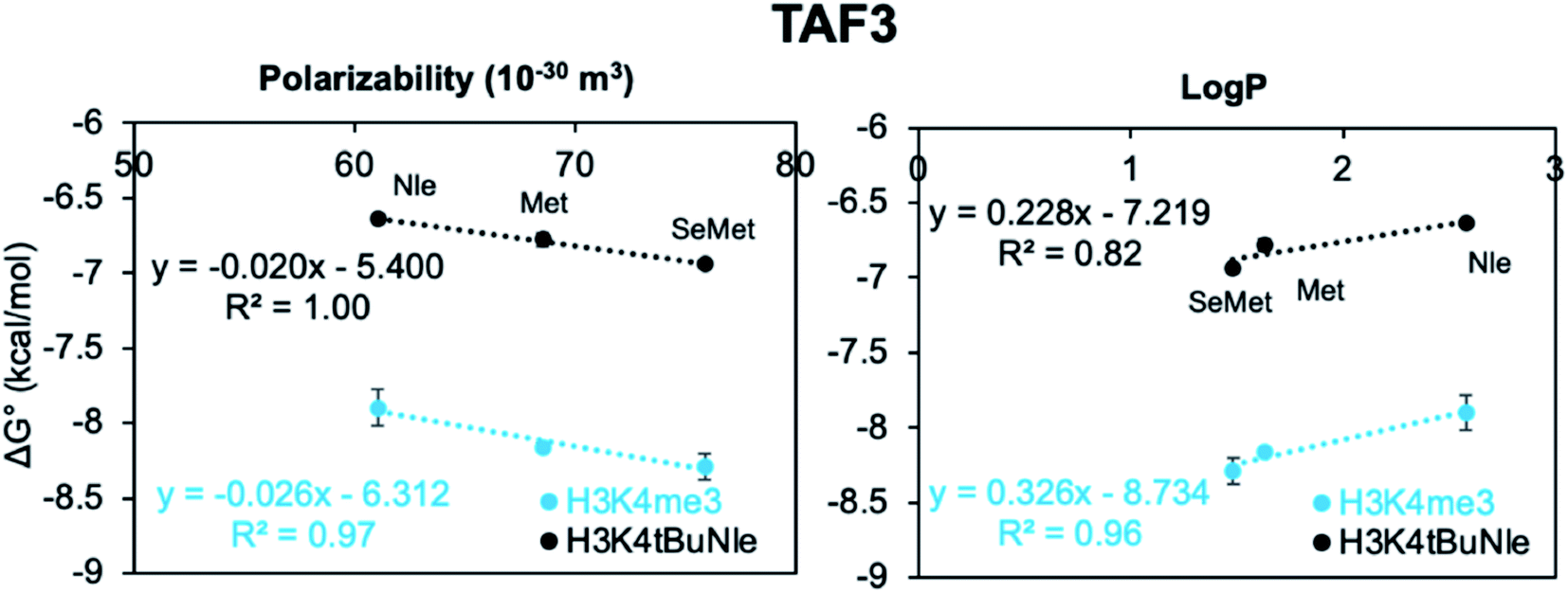
To evaluate the contributions of the different factors that may participate in binding, including dispersion forces and the hydrophobic effect, the free energy of binding 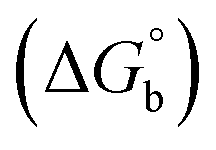 was plotted against polarizability, which is a measure of dispersion, and logP, which is a measure of hydrophobicity, to assess any correlations and identify possible linear free energy relationships (LFERs, Fig. 4). In general, the LFERs for TAF3 binding show better correlations than those for DIDO1. This is largely due to Met being the most favorable in DIDO1, but qualitatively, the trends are similar between the two. TAF3 shows a positive correlation with polarizability, but shows a negative correlation with logP (Fig. 4). This is consistent with previous reports that noncovalent interactions involving sulfur are driven by dispersion forces and that hydrophobic interactions do not dominate in this case.32–37,39–41
was plotted against polarizability, which is a measure of dispersion, and logP, which is a measure of hydrophobicity, to assess any correlations and identify possible linear free energy relationships (LFERs, Fig. 4). In general, the LFERs for TAF3 binding show better correlations than those for DIDO1. This is largely due to Met being the most favorable in DIDO1, but qualitatively, the trends are similar between the two. TAF3 shows a positive correlation with polarizability, but shows a negative correlation with logP (Fig. 4). This is consistent with previous reports that noncovalent interactions involving sulfur are driven by dispersion forces and that hydrophobic interactions do not dominate in this case.32–37,39–41
 | ||
| Fig. 4 LFERs for TAF3 (cyan) and DIDO1 (blue) binding to H3K4me3. | ||
DIDO1 has a weaker correlation with polarizability, and like TAF3, a negative correlation with logP, once again suggesting that the hydrophobic effect is not the major contributor. The correlation with polarizability suggests that for the Nle, Met, and SeMet mutants, dispersion contributes to binding, but does not directly address whether the charge on Kme3 is also important. Thus, to investigate the contribution of electrostatic interactions directly, we synthesized a neutral but equally polarizable analog of Kme3, tBuNle, and measured binding to DIDO1 and TAF3.
TAF3 is selective for Kme3 over tBuNle but DIDO1 and BPTF Y23M are unselective
Binding studies were performed using the neutral analog of Kme3, tBuNle,52,56 to determine the importance of the positive charge to the DIDO1 and TAF3 interactions with respect to mutation at the methionine position. Our group has previously developed tBuNle to show in a chromodomain reader protein that the replacement of Kme3 with tBuNle weakens binding by 2.2 kcal mol−1.56 Mecinović and coworkers have also previously shown that TAF3 binds H3K4tBuNle 2 kcal mol−1 less favorably than it binds H3K4me3.52 We confirmed that the neutral analog bound to TAF3 ten-fold weaker, and was less enthalpically favorable, consistent with a loss of cation–π interactions with the aromatic rings and any electrostatic component to the Met–ammonium interaction (Fig. 5). Prior to this report, binding to a neutral analog of Kme3 had not been investigated in DIDO1. Surprisingly, we found that the overall affinity for DIDO1 is within error of Kme3 and complete enthalpy–entropy compensation is observed (Fig. 5). The enthalpy–entropy compensation suggests different mechanisms for binding to the two ligands: cation–π interactions contribute to enthalpic binding Kme3, whereas tBuNle binding is driven by the hydrophobic effect, which is entropy-driven. A similar change in driving force has been observed in a β-hairpin model system that studied the Trp–Kme3 vs. Trp–tBuNle interactions.56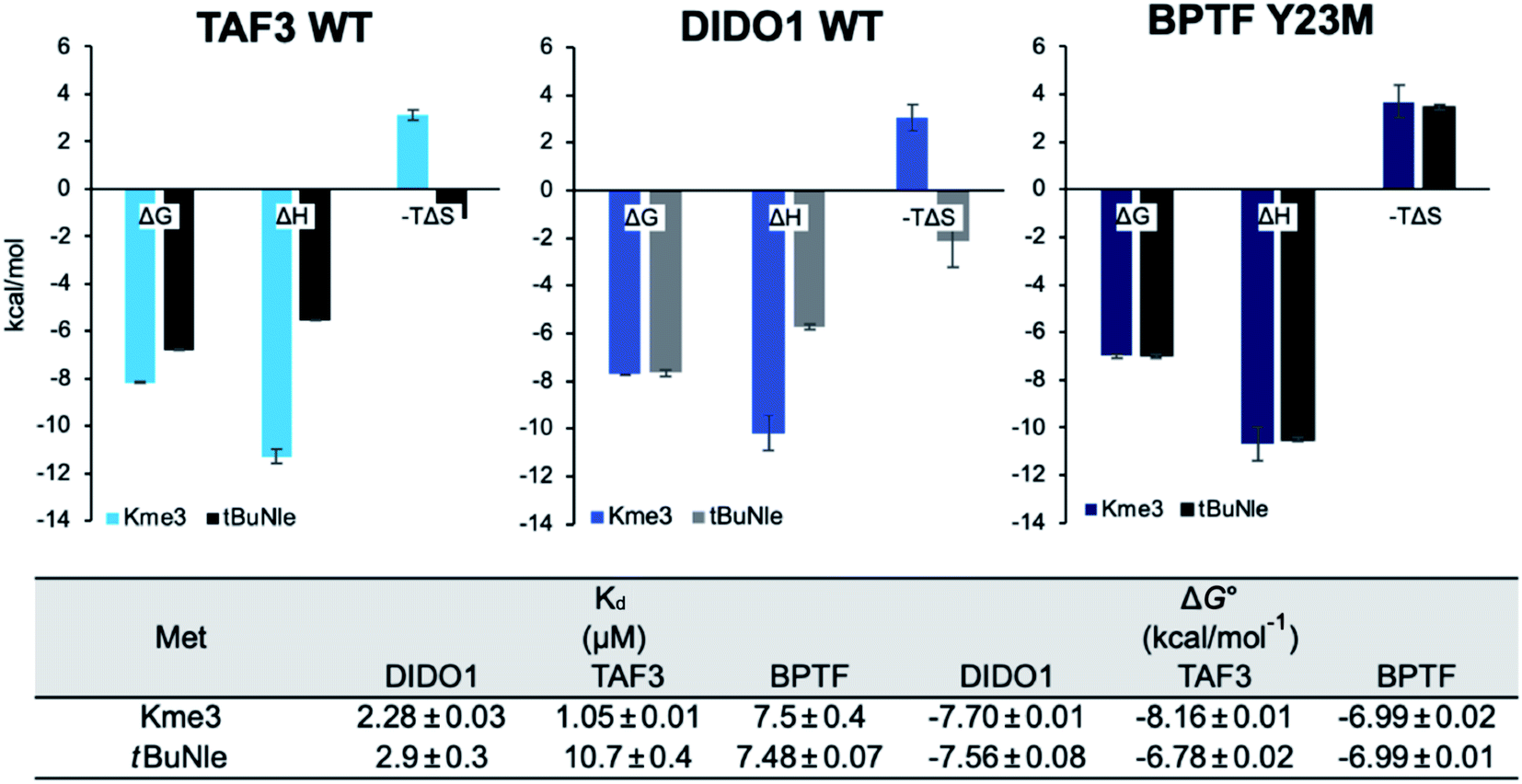 | ||
| Fig. 5 Thermodynamic consequences of DIDO1, TAF3, and BPTF Y23M binding H3K4me3 and H3K4tBuNle measured by ITC. Experiments were performed by titrating histone peptide (500 μM to 1 mM) into TAF3 and DIDO1 wild type domains (50–200 μM) in 50 mM sodium phosphate, pH 7.4, 150 mM NaCl, 2 mM TCEP at 25 °C. Error was determined as standard error of the mean. | ||
Like DIDO1, binding to the Y23M mutation in BPTF with the H3K4tBuNle ligand showed no loss in affinity between the two histone peptides, nor was there any change in thermodynamic driving force for the interaction (Fig. 5 and Table S6†). Mecinović and coworkers measured H3K4tBuNle binding to BPTF wild-type and found a less than two-fold difference in affinity compared to H3K4me3 (Table S6†).52 Thus, the Y23M mutation results in loss of selectivity for the charged residue, consistent with dispersion forces as the primary interaction between Met and the ligand. The lack of enthalpy–entropy compensation in BPTF Y23M demonstrates a different mechanism of binding from DIDO1 that is equally enthalpically favorable regardless of binding H3K4me3 and H3K4tBuNle.
Investigating the role of Met in achieving charge selectivity in TAF3
As TAF3 is selective for H3K4me3 over H3K4tBuNle and structural studies demonstrate that there is no change in the distance or geometry of TAF3 M882 to the bound state of either Kme3 or tBuNle (Fig. S16†),52 binding to the Met isosteres Nle and SeMet were investigated in this domain to further describe the influence of an electrostatic interaction specifically. The effects of the Met isosteres in TAF3 exhibit similar trends in affinity as binding to Kme3 (Table 3), with weaker binding to Nle and stronger binding to SeMet.| TAF3 mutationa | K d (μM) | ΔG° (kcal mol−1) | ΔΔG° (kcal mol−1) | |||
|---|---|---|---|---|---|---|
| tBuNle | Kme3 | tBuNle | Kme3 | tBuNle | Kme3 | |
| a Experiments were performed by titrating H3K4tBuNle peptide (500 μM to 1 mM) into TAF3 mutants (50–200 μM) in 50 mM sodium phosphate, pH 7.4, 150 mM NaCl, 2 mM TCEP at 25 °C. Error was determined as standard error of the mean. | ||||||
| Met | 10.7 ± 0.7 | 1.05 ± 0.01 | −6.78 ± 0.02 | −8.16 ± 0.01 | — | — |
| Nle | 13.7 ± 0.3 | 1.7 ± 0.2 | −6.64 ± 0.01 | −7.90 ± 0.06 | 0.15 ± 0.04 | 0.3 ± 0.2 |
| SeMet | 8.3 ± 0.2 | 0.84 ± 0.07 | −6.94 ± 0.01 | −8.29 ± 0.05 | −0.15 ± 0.04 | −0.1 ± 0.2 |
Analysis of the LFERs for tBuNle binding to TAF3 demonstrates that the trends are the same as with Kme3, showing positive correlations with 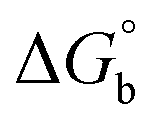 and polarizability but not logP (Fig. 6). It is surprising that logP is anti-correlative when binding to the neutral tBuNle analog, which has been previously shown to participate in hydrophobically driven interactions in other systems.56 The positive correlation with polarizability suggests that dispersion forces are also the main driving force for binding H3K4tBuNle (Fig. 6). Given that the polarizabilities for Kme3 and tBuNle are very similar (45.94 and 46.47 Å3, respectively), it is reasonable that the LFERs are not significantly different. The similar slopes of LFERs for TAF3 binding to H3K4me3 and H3K4tBuNle (Fig. 6) indicate that the magnitude of the Met interaction is nearly the same despite the loss of the charge on the ligand, thus the electrostatic contribution to the interaction is negligible. Moreover, these similarities are in agreement with the loss of charge selectivity that results from the Tyr-to-Met mutation in BPTF.
and polarizability but not logP (Fig. 6). It is surprising that logP is anti-correlative when binding to the neutral tBuNle analog, which has been previously shown to participate in hydrophobically driven interactions in other systems.56 The positive correlation with polarizability suggests that dispersion forces are also the main driving force for binding H3K4tBuNle (Fig. 6). Given that the polarizabilities for Kme3 and tBuNle are very similar (45.94 and 46.47 Å3, respectively), it is reasonable that the LFERs are not significantly different. The similar slopes of LFERs for TAF3 binding to H3K4me3 and H3K4tBuNle (Fig. 6) indicate that the magnitude of the Met interaction is nearly the same despite the loss of the charge on the ligand, thus the electrostatic contribution to the interaction is negligible. Moreover, these similarities are in agreement with the loss of charge selectivity that results from the Tyr-to-Met mutation in BPTF.
 | ||
| Fig. 6 LFERs for TAF3 comparing H3K4me3 (blue) to H3K4tBuNle (black). | ||
Conclusions
Using three PHD domains that recognize Kme3, we have investigated the contribution of the thioether–alkylammonium interaction to binding. Our approach of systematically varying the properties of a single residue within the aromatic cage of reader proteins55 allows us to discretely tune inherently weak noncovalent interactions to investigate their role in binding. These studies demonstrate that Met plays a favorable role in binding to Kme3 rather than simply contributing to protein structure. Additionally, LFERs indicate that the binding interaction between Met and Kme3 is driven primarily by dispersion forces and exhibits a negative correlation with hydrophobicity. Moreover, the correlation with polarizability holds true regardless of the charge on the ligand, suggesting there is not a unique sulfur–cation electrostatic interaction contributing to binding. In the context of sulfur based noncovalent interactions as a whole, our results correlate well with the literature precedent that Met largely interacts dispersively due to the highly polarizable sulfur atom in protein folding and protein–protein interactions.32–37,39–41In our investigation of the role of the ligand in the interaction with Met, we discovered a difference in the contribution of the charge to overall binding. Whereas TAF3 is selective for Kme3, DIDO1 is not, and instead demonstrates enthalpy–entropy compensation in binding tBuNle versus Kme3. Given dispersion driven interactions by Met are conserved between the two domains, our results suggest the change in mechanism from enthalpically to entropically favorable binding in DIDO1 must arise from interactions with the remainder of the aromatic cage. The enthalpy–entropy compensation observed in DIDO1 is consistent with a change in mechanism in binding from cation–π-driven to the hydrophobic effect between tBuNle and the aromatic cage. This same phenomenon has been observed previously in a β-hairpin peptide in the comparison of a Kme3 versus tBuNle interaction with a cross-strand Trp residue.56 Additionally, while BPTF, with a 4-residue aromatic cage, exhibits a modest 2-fold selective for Kme3,52 the BPTF Y23M mutant is no longer selective for Kme3 over tBuNle, nor is there any change in thermodynamic driving force for the interaction (Fig. 5 and Table S6†). The differences in selectivity for Kme3 over tBuNle between TAF3 and DIDO1, and BPTF Y23M were unexpected, and suggest a more complicated mode of binding than the cation–π mechanism that is generally assumed. Additional studies are underway to gain insight into these differences in selectivity.
Author contributions
M. L. W. and K. I. A. conceived the idea. M. L. W. directed the project. K. I. A. performed the experiments. The manuscript was written and edited by K. I. A. and M. L. W.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This work was supported by the National Institutes of Health, National Institute of General Medical Sciences (GM118499). KIA was supported in part by a Burroughs Wellcome Fellowship.References
- T. Kouzarides, Chromatin Modifications and Their Function, Cell, 2007, 128, 693–705 CrossRef CAS PubMed.
- B. C. Smith and J. M. Denu, Chemical mechanisms of histone lysine and arginine modifications, Biochim. Biophys. Acta, Gene Regul. Mech., 2009, 1789, 45–57 CrossRef CAS PubMed.
- E. L. Greer and Y. Shi, Histone methylation: a dynamic mark in health, disease and inheritance, Nat. Rev. Genet., 2012, 13, 343–357 CrossRef CAS PubMed.
- K. Hyun, J. Jeon, K. Park and J. Kim, Writing, erasing and reading histone lysine methylations, Exp. Mol. Med., 2017, 49, 324 CrossRef.
- M. A. Dawson and T. Kouzarides, Cancer Epigenetics: From Mechanism to Therapy, Cell, 2012, 150, 12–27 CrossRef CAS PubMed.
- E. A. Mazzio and K. F. A. Soliman, Basic concepts of epigenetics: impact of environmental signals on gene expression, Epigenetics, 2012, 7, 119–130 CrossRef CAS PubMed.
- C. D. Allis and T. Jenuwein, The molecular hallmarks of epigenetic control, Nat. Rev. Genet., 2016, 17, 487–500 CrossRef CAS PubMed.
- L. I. James and S. V. Frye, Targeting Chromatin Readers, Clin. Pharmacol. Ther., 2013, 93, 312–314 CrossRef CAS PubMed.
- K. Helin and D. Dhanak, Chromatin proteins and modifications as drug targets, Nature, 2013, 502, 480–488 CrossRef CAS PubMed.
- S. Maurer-Stroh, N. J. Dickens, L. Hughes-Davies, T. Kouzarides, F. Eisenhaber and C. P. Ponting, The Tudor domain ‘Royal Family’: Tudor, plant Agenet, Chromo, PWWP and MBT domains, Trends Biochem. Sci., 2003, 28, 69–74 CrossRef CAS.
- S. D. Taverna, H. Li, A. J. Ruthenburg, C. D. Allis and D. J. Patel, How chromatin-binding modules interpret histone modifications: lessons from professional pocket pickers, Nat. Struct. Mol. Biol., 2007, 14, 1025–1040 CrossRef CAS.
- W. Fischle, H. Franz, S. A. Jacobs, C. D. Allis and S. Khorasanizadeh, Specificity of the Chromodomain Y Chromosome Family of Chromodomains for Lysine-methylated ARK(S/T) Motifs, J. Biol. Chem., 2008, 283, 19626–19635 CrossRef CAS.
- R. Sanchez and M.-M. Zhou, The PHD finger: a versatile epigenome reader, Trends Biochem. Sci., 2011, 36, 364–372 CAS.
- C. A. Musselman, M.-E. Lalonde, J. Côté and T. G. Kutateladze, Perceiving the epigenetic landscape through histone readers, Nat. Struct. Mol. Biol., 2012, 19, 1218–1227 CrossRef CAS.
- A. W. Wilkinson and O. Gozani, Histone-binding domains: Strategies for discovery and characterization, Biochim. Biophys. Acta, Gene Regul. Mech., 2014, 1839, 669–675 CrossRef CAS PubMed.
- F. H. Andrews, B. D. Strahl and T. G. Kutateladze, Insights into newly discovered marks and readers of epigenetic information, Nat. Chem. Biol., 2016, 12, 662–668 CrossRef CAS PubMed.
- J. Gatchalian, M. C. Kingsley, S. D. Moslet, R. D. Rosas Ospina and T. G. Kutateladze, An aromatic cage is required but not sufficient for binding of Tudor domains of the polycomblike protein family to H3K36me3, Epigenetics, 2015, 10(6), 467–473 CrossRef PubMed.
- J. C. Ma and D. A. Dougherty, The Cation–π Interaction, Chem. Rev., 1997, 97, 1303–1324 CrossRef CAS PubMed.
- P. V. Peña, F. Davrazou, X. Shi, K. L. Walter, V. V. Verkhusha, R. Zhao and T. G. Kutateladze, Molecular mechanism of histone H3K4me3 recognition by plant homeodomain of ING2, Nature, 2006, 442, 100–103 CrossRef PubMed.
- S. Ramón-Maiques, A. J. Kuo, D. Carney, A. G. W Matthews, M. A. Oettinger, O. Gozani and W. Yang, The plant homeodomain finger of RAG2 recognizes histone H3 methylated at both lysine-4 and arginine-2, Proc. Natl. Acad. Sci. U. S. A., 2007, 104, 18993–18998 CrossRef PubMed.
- M. Vermeulen, K. W. Mulder, S. Denissov, W. W. M. P. Pijnappel, F. M. A. van Schaik, R. A. Varier, M. P. A. Baltissen, H. G. Stunnenberg, M. Mann and H. T. M. Timmers, Selective anchoring of TFIID to nucleosomes by trimethylation of histone H3 lysine 4, Cell, 2007, 131, 58–69 CrossRef CAS PubMed.
- S. Ramón-Maiques, A. J. Kuo, D. Carney, A. G. W. Matthews, M. A. Oettinger, O. Gozani and W. Yang, The plant homeodomain finger of RAG2 recognizes histone H3 methylated at both lysine-4 and arginine-2, Proc. Natl. Acad. Sci. U. S. A., 2007, 104, 18993–18998 CrossRef PubMed.
- C. A. Musselman and T. G. Kutateladze, PHD fingers: epigenetic effectors and potential drug targets, Mol. Interventions, 2009, 9, 314–323 CrossRef CAS PubMed.
- J. Gatchalian, A. Fütterer, S. B. Rothbart, Q. Tong, H. Rincon-Arano, A. Sánchez de Diego, M. Groudine, B. D. Strahl, C. Martínez-A, K. H. M. van Wely and T. G. Kutateladze, Dido3 PHD Modulates Cell Differentiation and Division, Cell Rep., 2013, 4, 148–158 CrossRef CAS PubMed.
- K. Kinkelin, G. G. Wozniak, S. B. Rothbart, M. Lidschreiber, B. D. Strahl, P. Cramer and R. Huber, Structures of RNA polymerase II complexes with Bye1, a chromatin-binding PHF3/DIDO homologue, Proc. Natl. Acad. Sci. U. S. A., 2013, 110, 15277–15282 CrossRef CAS PubMed.
- A. H. Tencer, J. Gatchalian, B. J. Klein, A. Khan, Y. Zhang, B. D. Strahl, K. H. M. van Wely and T. G. Kutateladze, A Unique pH-Dependent Recognition of Methylated Histone H3K4 by PPS and DIDO, Structure, 2017, 25, 1530–1539.e3 CrossRef CAS PubMed.
- H. van Ingen, F. M. A. van Schaik, H. Wienk, J. Ballering, H. Rehmann, A. C. Dechesne, J. A. W. Kruijzer, R. M. J. Liskamp, H. T. M. Timmers and R. Boelens, Structural insight into the recognition of the H3K4me3 mark by the TFIID subunit TAF3, Structure, 2008, 16, 1245–1256 CrossRef CAS PubMed.
- K. Hyun, J. Jeon, K. Park and J. Kim, Writing, erasing and reading histone lysine methylations, Exp. Mol. Med., 2017, 49, 324 CrossRef PubMed.
- C. C. Valley, A. Cembran, J. D. Perlmutter, A. K. Lewis, N. P. Labello, J. Gao and J. N. Sachs, The methionine-aromatic motif plays a unique role in stabilizing protein structure, J. Biol. Chem., 2012, 287, 34979–34991 CrossRef CAS PubMed.
- J. Hwang, P. Li, M. D. Smith, C. E. Warden, D. A. Sirianni, E. C. Vik, J. M. Maier, C. J. Yehl, C. David Sherrill and K. D. Shimizu, Tipping the Balance between S–π and O–π Interactions, J. Am. Chem. Soc., 2018, 20, 27 Search PubMed.
- K. N.-M. Daeffler, H. A. Lester and D. A. Dougherty, Functionally Important Aromatic–Aromatic and Sulfur–π Interactions in the D2 Dopamine Receptor, J. Am. Chem. Soc., 2012, 134(36), 14890–14896 CrossRef CAS PubMed.
- R. Newberry and R. T. Raines, Secondary Forces in Protein Folding, ACS Chem. Biol., 2019, 14, 1677–1686 CrossRef CAS.
- J. Pranata, Sulfur–Aromatic Interactions: A Computational Study of the Dimethyl Sulfide–Benzene Complex, Bioorg. Chem., 1997, 25, 213–219 CrossRef CAS.
- R. J. Zauhar, C. L. Colbert, R. S. Morgan and W. J. Welsh, Evidence for a strong sulfur–aromatic interaction derived from crystallographic data, Biopolymers, 2000, 53, 233–248 CrossRef CAS PubMed.
- C. D. Tatko and M. L. Waters, Investigation of the nature of the methionine–π interaction in β-hairpin peptide model systems, Protein Sci., 2004, 13, 2515–2522 CrossRef CAS PubMed.
- H. S. Biswal and S. Wategaonkar, Sulfur, Not Too Far Behind O, N, and C: SH···π Hydrogen Bond, J. Phys. Chem. A, 2009, 113, 12774–12782 CrossRef CAS.
- B. R. Beno, K. S. Yeung, M. D. Bartberger, L. D. Pennington and N. A. Meanwell, A Survey of the Role of Noncovalent Sulfur Interactions in Drug Design, J. Med. Chem., 2015, 58, 4383–4438 CrossRef CAS PubMed.
- J. C. Gomez-Tamayo, A. Cordomı, M. Olivella, E. Mayol, D. Fourmy and L. Pardo, Analysis of the interactions of sulfur-containing amino acids in membrane proteins, Protein Sci., 2016, 25, 1517–1524 CrossRef CAS PubMed.
- A. R. Viguera and L. Serrano, Side-Chain Interactions between Sulfur-Containing Amino Acids and Phenylalanine in a-Helices The side-chain–side-chain interaction between Phe residues and sulfur-containing residues, Biochemistry, 1995, 34, 8771–8779 CrossRef CAS PubMed.
- E. A. Meyer, R. K. Castellano and F. Diederich, Interactions with Aromatic Rings in Chemical and Biological Recognition, Angew. Chem., Int. Ed., 2003, 42, 1210–1250 CrossRef CAS PubMed.
- L. M. Salonen, M. Ellermann and F. Diederich, Aromatic Rings in Chemical and Biological Recognition: Energetics and Structures, Angew. Chem., Int. Ed., 2011, 50, 4808–4842 CrossRef CAS PubMed.
- H. Duewel, E. Daub, V. Robinson and J. F. Honek, Incorporation of Trifluoromethionine into a Phage Lysozyme: Implications and a New Marker for Use in Protein 19 F NMR, Biochemistry, 1997, 36, 3404–3416 CrossRef CAS PubMed.
- M. D. Vaughan, P. Cleve, V. Robinson, H. S. Duewel and J. F. Honek, Difluoromethionine as a Novel 19 F NMR Structural Probe for Internal Amino Acid Packing in Proteins, J. Am. Chem. Soc., 1999, 121, 8475–8478 CrossRef CAS.
- D. K. Garner, M. D. Vaughan, H. J. Hwang, M. G. Savelieff, S. M. Berry, J. F. Honek and Y. Lu, Reduction Potential Tuning of the Blue Copper Center in Pseudomonas aeruginosa Azurin by the Axial Methionine as Probed by Unnatural Amino Acids, J. Am. Chem. Soc., 2006, 128, 15608–15617 CrossRef CAS PubMed.
- J. F. Honek, Effects of Fluorination on the Bioorganic Properties of Methionine, in Fluorine in Medicinal Chemistry and Chemical Biology, 2009 Search PubMed.
- M. D. Vaughan, Z. Su, E. Daub and J. F. Honek, Intriguing cellular processing of a fluorinated amino acid during protein biosynthesis in Escherichia coli, Org. Biomol. Chem., 2016, 14, 8942–8946 RSC.
- J. C. M. Van Hest, K. L. Kiick and D. A. Tirrell, Efficient Incorporation of Unsaturated Methionine Analogues into Proteins in Vivo, J. Am. Chem. Soc., 2000, 122, 1282–1288 CrossRef CAS.
- Y. Liu, H. Kim, J. Liang, W. Lu, B. Ouyang, D. Liu and Z. Songyang, The Death-Inducer Obliterator 1 (Dido1) Gene Regulates Embryonic Stem Cell Self-renewal, J. Biol. Chem., 2013, 289, 4778–4786 CrossRef.
- O. Bereczki, Z. Ujfaludi, N. Pardi, Z. Nagy, L. Tora, I. M. Boros and E. Balint, TATA binding protein associated factor 3 (TAF3) interacts with p53 and inhibits its function, BMC Mol. Biol., 2008, 9, 57 CrossRef PubMed.
- B. Pieters, E. Meulenbroeks, R. Belle and J. Mecinović, The Role of Electrostatic Interactions in Binding of Histone H3K4me2/3 to the Sgf29 Tandem Tudor Domain, PLoS One, 2015, 10, 1–15 Search PubMed.
- C. He, N. Liu, D. Xie, Y. Liu, Y. Xiao and F. Li, Structural basis for histone H3K4me3 recognition by the N-terminal domain of the PHD finger protein Spp1, Biochem. J., 2019, 476, 1957–1973 CrossRef CAS PubMed.
- J. J. A. G. Kamps, J. Huang, J. Poater, C. Xu, B. J. G. E. Pieters, A. Dong, J. Min, W. Sherman, T. Beuming, F. Matthias Bickelhaupt, H. Li and J. Mecinović, Chemical basis for the recognition of trimethyllysine by epigenetic reader proteins, Nat. Commun., 2015, 6, 8911 CrossRef CAS PubMed.
- C. M. Santiveri, M. F. García-Mayoral, J. M. Pérez-Cañadillas and M. Á. Jiménez, NMR structure note: PHD domain from death inducer obliterator protein and its interaction with H3K4me3, J. Biomol. NMR, 2013, 56, 183–190 CrossRef CAS PubMed.
- S. Hovmoller, T. Zhou and T. Ohlson, Conformations of amino acids in proteins, Acta Crystallogr., Sect. D: Biol. Crystallogr., 2002, 58, 768–776 CrossRef PubMed.
- S. A. Baril, A. L. Koenig, M. W. Krone, K. I. Albanese, C. Q. He, G. Y. Lee, K. N. Houk, M. L. Waters and E. M. Brustad, Investigation of Trimethyllysine Binding by the HP1 Chromodomain via Unnatural Amino Acid Mutagenesis, J. Am. Chem. Soc., 2017, 139, 17253–17256 CrossRef CAS PubMed.
- R. M. Hughes, K. R. Wiggins, S. Khorasanizadeh and M. L. Waters, Recognition of trimethyllysine by a chromodomain is not driven by the hydrophobic effect, Proc. Natl. Acad. Sci. U. S. A., 2007, 104, 11184–11188 CrossRef CAS PubMed.
- H. Li, S. Ilin, W. Wang, E. M. Duncan, J. Wysocka, C. D. Allis and D. J. Patel, Molecular basis for site-specific read-out of histone H3K4me3 by the BPTF PHD finger of NURF, Nature, 2006, 442, 91–95 CrossRef CAS PubMed.
- J. C. Aledo, Methionine in proteins: The Cinderella of the proteinogenic amino acids, Protein Sci., 2019, 28, 1785–1796 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d1sc02175c |
| This journal is © The Royal Society of Chemistry 2021 |