
Tuning the affinity of probes with transmembrane proteins by constructing peptide-conjugated cis/trans isomers based on molecular scaffolds†
Jing-Jing
Hu
a,
Juliang
Yang
a,
Yiheng
Liu
a,
Guangwen
Lu
a,
Zujin
Zhao
 b,
Fan
Xia
a and
Xiaoding
Lou
*a
b,
Fan
Xia
a and
Xiaoding
Lou
*a
aState Key Laboratory of Biogeology and Environmental Geology, Faculty Materials Science and Chemistry, China University of Geosciences, Wuhan 430078, China. E-mail: louxiaoding@cug.edu.cn
bDepartment State Key Laboratory of Luminescent Materials and Devices, Guangdong Provincial Key Laboratory of Luminescence from Molecular Aggregates, South China University of Technology, Guangzhou 510640, China
First published on 23rd October 2024
Abstract
For protein analysis, the current peptide-based probes rely almost on the specific recognition of the protein while neglecting the potential influence of the environment near the protein. Herein, we propose that to achieve high recognition of transmembrane protein integrin αvβ3, the interactions from the membrane substrate could be helpful. Moreover, to guarantee the additive effect of different interactions, the cis and trans isomers of peptide-based probes are distinguished. In detail, we synthesized the peptide-conjugated cis/trans isomers (cis-RTP and trans-RTP) by modifying the Arg-Gly-Asp (RGD)-targeting peptide and palmitic acid-conjugated Arg-Arg-Arg-Arg (Pal-RRRR) peptide to the two ends of the molecular scaffold-tetraphenylethene derivative. Due to the difference in spatial structure, isothermal titration calorimetry and simulation experiments demonstrated that cis-RTP can bind more stably to integrin αvβ3 than trans-RTP. As a result, cis-RTP has shown more excellent properties in inhibiting cell migration and killing cells by regulating actin and extracellular signal-regulated kinase. Unlike the existing probe design for protein, this study provides a concept of microenvironment-helpful recognition and a promising strategy of cis/trans isomers to modulate the interaction between proteins and probes.
Introduction
Transmembrane proteins, constituting about 30% of the mammalian proteome, participate in a series of significant physiological activities, like mediating metabolism, controlling signals, transporting molecules, and others required for cellular life.1–3 Therefore, systematically investigating and gaining in-depth understanding of transmembrane proteins are significant for elucidating the molecular basis of biological processes, disease developments, and molecular mechanisms of disease treatments.4–6 Generally, their analysis relies on the recognition and interaction between transmembrane proteins and probes, like antibodies, aptamers, and peptides.7–12 For instance, Tan et al. developed a technology of introducing artificial nucleotides with specific chemical groups to the cell systematic evolution of ligands by exponential enrichment (SELEX) process, thereby obtaining aptamers with the ability to regulate transmembrane protein activity.13 Utilizing the method of mirror-image phage display, Gao et al. obtained the D-peptide with hydrolysis-resistant capability and high affinity to bind to the transmembrane protein programmed cell death protein 1 (PD-1), thus blocking the original PD-1/PD-L1 pathway to achieve tumor immunotherapy.14 Among these recognition probes, peptides with endogenous bioactivity, higher editability and biocompatibility have unique advantages in precise molecular targeting and could be used as powerful identification tools.15–26 However, current acquisition methods for peptide probes mainly rely on molecular screening or molecular evolution methods for protein targets alone, neglecting the influence of the environment.In fact, transmembrane proteins commonly do not exist in isolation but are in the complex membrane environment and specific substrate, most obviously the phospholipid bilayer.27 It has been reported that unlike cytoplasmic proteins, transmembrane proteins are solvated by the complex lipid matrix, unavoidably affecting protein structure, dynamics, and function in diverse ways. Apart from substrates, lipids can also serve as regulatory co-factors for transmembrane proteins.28 Motivated by these studies, we wonder if we could fully use the membrane environment to propose a new strategy for improving the recognition of membrane proteins, which is less reported as far as we know.
In this proof-of-concept study, a typical and common transmembrane protein integrin αvβ3 was selected as the target. It can not only activate intracellular signalling by binding to ligands but also be the main receptor for cell adhesion and regulate cell migration.29–33 Herein, we constructed the peptide-conjugated probes by utilizing the classical integrin αvβ3-targeting peptide Arg-Gly-Asp (RGD) as a specific recognition unit and designing aggregation-induced emission luminogen (AIEgen)34,35 tetraphenylethene with two different functional groups (T-MY) as a fluorescent molecular scaffold, especially equipping with an additional hydrophobic palmitic acid-conjugated positively charged Arg-Arg-Arg-Arg peptide (Pal-RRRR), which could interact with the surrounding membrane substrate through two weak interactions (electrostatic interaction and hydrophobic interaction). As shown in Scheme 1, when RGD and Pal-RRRR were conjugated to the same-side benzene rings on the carbon–carbon double bond through the alkynyl–azide “click” reaction and thiol-Michael addition reaction, cis-RTP could be obtained. As to the two sides of the carbon–carbon double bond, trans-RTP could be constructed. By distinguishing the RTP isomers, the interactions ascribing to the membrane could be guaranteed to be devoted to the affinity of the transmembrane protein. After successful differentiation of the cis-RTP and trans-RTP probes, their affinities to integrin αvβ3 on the cell membrane were compared. Additionally, the diverse cell behaviours, including cell migration and cell growth induced by different probes, were investigated. This study proposed a new factor that can be considered when designing a protein analysis probe and provided a novel strategy to modulate the affinity with proteins by distinguishing the isomers.
 | ||
| Scheme 1 Chemical structures of cis-RTP/trans-RTP and their performance differences toward integrin αvβ3 on cell membrane. The cis-RTP interacts more stably with cell membrane than trans-RTP. By interacting with integrin αvβ3, cis-RTP has shown more excellent properties in inhibiting cell migration and killing cells by regulating actin and ERK through intracellular proteins. | ||
Results and discussion
Separation and characterization of peptide-conjugated cis/trans isomers
RTP was synthesized through two-step chemical reactions, including a thiol-Michael addition reaction between Pal-RRRR and T-MY to obtain the TP probe, and then the alkynyl-azide “click” reaction to conjugate TP with RGD peptide (Schemes S1, S2 and Table S1, ESI†). There were two chromatographic peaks in the cis-RTP and trans-RTP isomers, and their retention times were 8.9 min and 12.2 min, respectively (Fig. 1A and Table S2, ESI†). Due to the different dipole moments, the molecular polarity of cis-RTP is larger than that of trans-RTP. Therefore, in reversed-phase high-performance liquid chromatography (HPLC), the retention time of the more polar substances is shorter than the retention time of the less polar substances. According to the relative relationship between molecular polarity and retention time, the retention time of cis-RTP is shorter than that of trans-RTP. The high-resolution mass spectra (HRMS) and 1H nuclear magnetic resonance (NMR) proved the chemical structural formula of cis-RTP and trans-RTP (Fig. S1–S5, ESI†). Moreover, it was found that cis-RTP and trans-RTP showed similar absorption spectra attributing to the existence of AIEgen T-MY (300–400 nm) (Fig. S6, ESI†), while the fluorescent (FL) intensity of cis-RTP was higher than that of trans-RTP visibly in the range of 450–700 nm (Fig. 1B). To explore the reason for the different fluorescence intensities of isomers, ground-state geometry optimization calculations were further performed. As shown in Fig. 1C, the volume of cis-RTP (14935.564 Å3) was significantly smaller than that of trans-RTP (20682.483 Å3), which resulted in a larger molecular steric hindrance to cis-RTP, thus leading to brighter fluorescence. Collectively, the above results demonstrated the successful synthesis and isolation of cis-RTP and trans-RTP. However, the general tool 1H–1H COSY NMR spectra for further proving the structures of cis-RTP and trans-RTP were indistinguishable (Fig. S7 and S8, ESI†), which might be ascribed to the peptide conjugation-induced complex molecular structure and large molecular weight of RTP.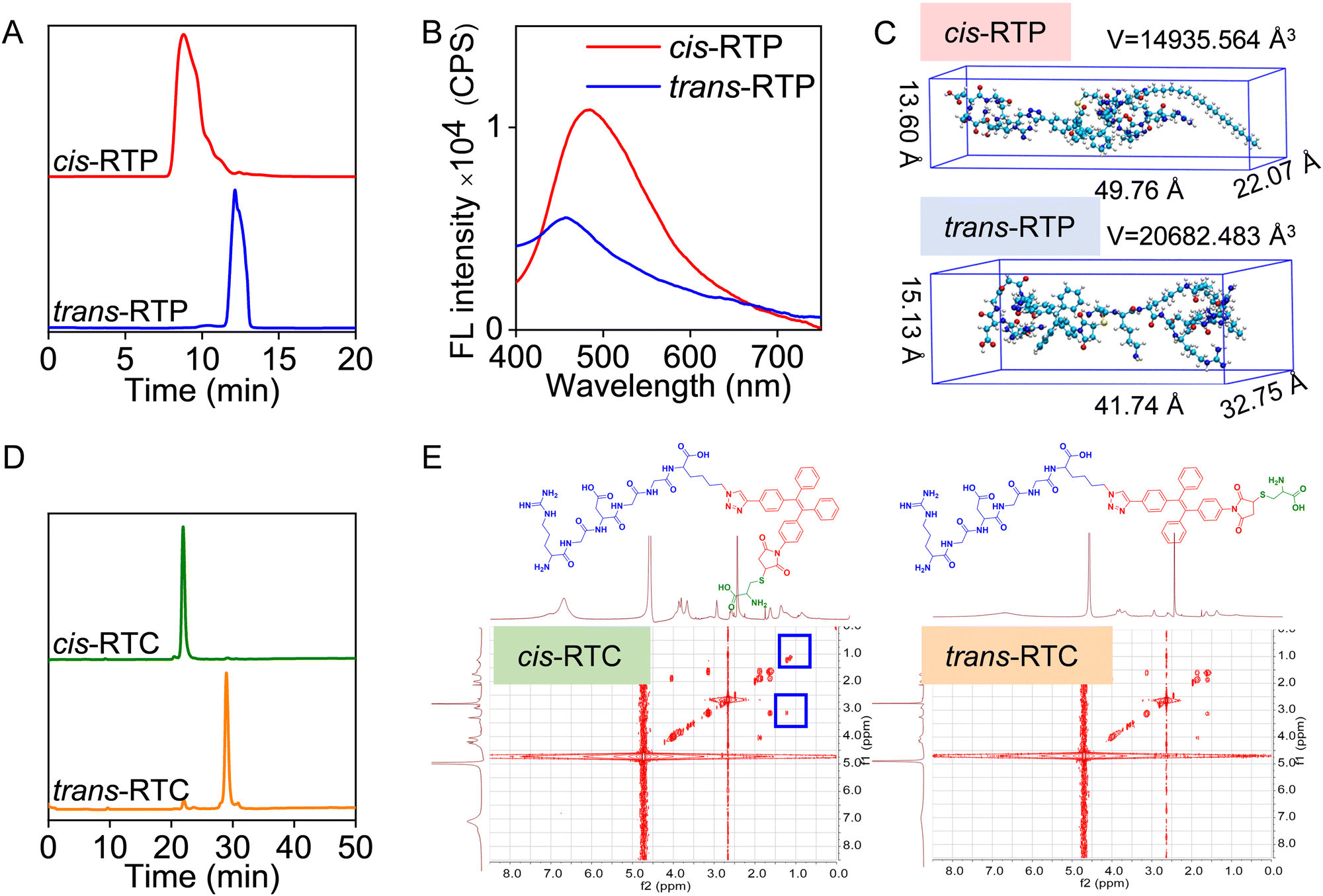 | ||
| Fig. 1 Separation and characterization of probes. (A) HPLC purification results of cis-RTP/trans-RTP. (B) FL spectra of cis-RTP/trans-RTP (10 μM) in DMSO/water mixture (v/v = 1/99). λex = 330 nm. (C) Ground-state geometry optimization calculation results of cis-RTP/trans-RTP. (D) HPLC purification results of cis-RTC/trans-RTC. (E) Chemical structural formulas and 1H–1H COSY NMR spectroscopies of cis-RTC/trans-RTC. | ||
In this case, to further confirm the successful synthesis of cis-RTP and trans-RTP, we simplified the structure of RTP and constructed another pair of isomers (cis-RTC and trans-RTC) by adopting the cystine instead of Pal-RRRR. Additionally, cis-RTC and trans-RTC were synthesized using the same two-step reactions. The results of HPLC separation showed that the retention times were 21.8 min and 29.1 min, respectively (Fig. 1D and Table S3, ESI†). According to the same analysis methods, we concluded that the probe at 21.8 min was cis-RTC and another one was trans-RTC. Mass spectra and 1H NMR spectra also demonstrated their successful synthesis (Fig. S9–S13, ESI†). The absorption and FL spectra also showed that they exhibited similar optical properties (Fig. S14, ESI†). Then, 1H–1H COSY NMR spectra were utilized to justify the judgment of the cis–trans structure of RTC. As shown in the 1H–1H COSY spectrum of cis-RTC (Fig. 1E), the resonance at δ = 1.2 ppm corresponded to cysteine protons (green part), and the resonance at δ = 3.1 ppm was due to RGD protons (blue part). This showed that cysteine and RGD were close in space, which is consistent with the steric configuration of cis-RTC. In contrast, trans-RTC did not have this phenomenon because the steric structures of its RGD and cysteine were not close. These data strongly proved our inference about cis-RTC/trans-RTC in the above. Because the relative polarity of cis-RTC/trans-RTC was consistent with that of cis-RTP/trans-RTP, our previous judgment on cis-RTP/trans-RTP was correct.
The binding ability of the peptide-conjugated cis/trans isomers to the integrin αvβ3 on the cell membrane
According to the intramolecular motion restriction mechanism of AIE, AIEgen-based probes could emit bright fluorescence after binding to the target.24 Thus, the specificity of probes bound to integrin was explored by fluorescence spectroscopy (Fig. 2A and Fig. S15, S16, ESI†). It was found that compared to treatment with other proteins, including fibroblast activation protein-α (FAP-α), phosphatase, bovine serum albumin (BSA) and matrix metalloproteinase 2 (MMP-2), cis/trans-RTP and cis/trans-RTC, exhibited significantly enhanced fluorescence after treatment with integrin αvβ3, suggesting that all the four probes containing RGD peptide could specifically bind to integrin αvβ3. Because integrin αvβ3 was highly expressed on the cell membrane of ovarian cancer cells (Fig. S17, ESI†), we extracted the cell membrane of ovarian cancer cell line SKOV3 to study the binding situation of cis-RTP/trans-RTP and cis-RTC/trans-RTC to the integrin αvβ3 on the membrane, as illustrated in Fig. 2B and Fig. S18 (ESI†). Compared to the trans-RTP or RTC isomers with a single interaction, the fluorescence intensity increase ratio of cis-RTP was much larger. This might be related to their different binding forces to the integrin αvβ3. To demonstrate this, the affinity of the probes to the extracted integrin αvβ3 on the membrane was further measured by isothermal titration calorimetry (ITC) experiments (Fig. 2C, D and Fig. S19, ESI†). The normalized binding constant indicated that cis-RTP was significantly higher than trans-RTP and cis-RTC/trans-RTC. Simultaneously, trans-RTP showed two binding constants, which were due to the influence of its spatial structure. The binding process of RGD to integrin αvβ3 and the binding process of Pal-RRRR to the cell membrane cannot be carried out simultaneously. In contrast, the spatial structure of cis-RTP allowed its RGD and Pal-RRRR to bind to the cell membrane simultaneously, thus leading to the strongest binding force (Ka = 1.0 × 107 M−1). Besides, the computational results demonstrated that the angles between RGD and Pal-RRRR to the central AIEgen were 116.02° and 172.67° for cis-RTP and trans-RTP, respectively, further proving the relatively close distance for two arms in cis-RTP, which would help to interact with integrin and cell membranes simultaneously (Fig. S20, ESI†). As to trans-RTP, it is hard for the two arms to bind to the integrin αvβ3 and insert into the cell membrane simultaneously, which may be due to the too-strong twisting force in the central structure (i.e., T-MY). These phenomena were also observed by simulating the probes with integrin αvβ3 on the cell membrane (Fig. 3). The binding energy of cis-RTP to the integrin αvβ3 on the cell membrane was calculated to be −672.637 kJ mol−1, and the binding energy of trans-RTP to that was −462.971 kJ mol−1, illustrating the stronger and more stable binding state of cis-RTP. These results demonstrate that owing to the difference in spatial structure, cis-RTP has a higher affinity with integrin αvβ3 on the cell membrane and can bind to it more stably compared with trans-RTP. | ||
| Fig. 2 Binding ability of the probe to the integrin αvβ3. (A) Plot of (I − I0)/I0versus different proteins, where I and I0 are the FL intensities at analyte concentrations of 100 and 0 μg mL−1. The proteins include integrin αvβ3, fibroblast activation protein-α (FAP-α), phosphatase, bovine serum albumin (BSA) and matrix metalloproteinase 2 (MMP-2). The concentration of the probes was 40 μM, λex = 330 nm. (B) Scheme of the fluorescence experiment and plot of (I30 − I0)/I0. I30 and I0 are the FL intensities at analyte times of 30 min and 0 min. (C) ITC results of cis-RTP in PBS with cell membrane at 37 °C. The concentration of the probes was 40 μM. (D) Binding constant calculation of cis-RTP/trans-RTP and cis-RTC/trans-RTC. Data are expressed as mean ± SD; **p < 0.01, ***p < 0.001. | ||
 | ||
| Fig. 3 Theoretical binding energy calculation results of (A) cis-RTP and (B) trans-RTP. Multicolor, blue, and gray represent the probe, integrin αvβ3, and phospholipid bilayer, respectively. | ||
Effects of peptide-conjugated cis/trans isomers on cell adhesion, migration and cell growth
Studies have shown that RGD-targeted integrin αvβ3 are not used only as targeting receptors but also regulate cell adhesion and migration, and activate intracellular signalling (Fig. 4A).36,37 Thus, we next evaluated whether the isomers with different affinities with integrin αvβ3 could induce the diverse living cell responses. Both confocal laser scanning microscope (CLSM) and 2.5 D imaging results showed that cis-RTP/trans-RTP involved in RGD peptides could target tumor cell membranes (Fig. 4B). Then, the effects of cis-RTP/trans-RTP and cis-RTC/trans-RTC on SKOV3 cell adhesion were assessed, as depicted in Fig. 4C, D and Fig. S21 (ESI†). Here, the cis-RTC/trans-RTC can be used as the control group, with two fewer interactions compared to RTP isomers. The quantitative results showed that the number of adherent cells in the cis-RTP-treated cells was significantly lower than that in the trans-RTP and cis-RTC/trans-RTC groups. Besides, the number of adherent cells in the other three groups was similar to that in the blank group, indicating that cis-RTP could reduce the adhesion behaviour of cells, while trans-RTP and cis-RTC/trans-RTC with lower affinity for integrin αvβ3 could not. Subsequently, the migration ability of SKOV3 cells treated with different probes was investigated through scratch-wound assay experiments (Fig. 4E, F and Fig. S22, ESI†). The ratio of scratched area at 24 h versus scratched area at 0 h was used to quantify the cell migration situation. The ratio of cis-RTP-treated cells was about 0.70, while that of the other groups was about 0.50 or less, suggesting that cis-RTP also greatly influenced the migratory behavior of cells. All the above results imply that despite the same peptide modules, there were obvious differences in the effects of the cis/trans structure of RTP on the adhesion and migration behavior of cells. Importantly, the effects of cis-RTP were much greater than those of trans-RTP, which is consistent with their differences in affinity to integrin αvβ3.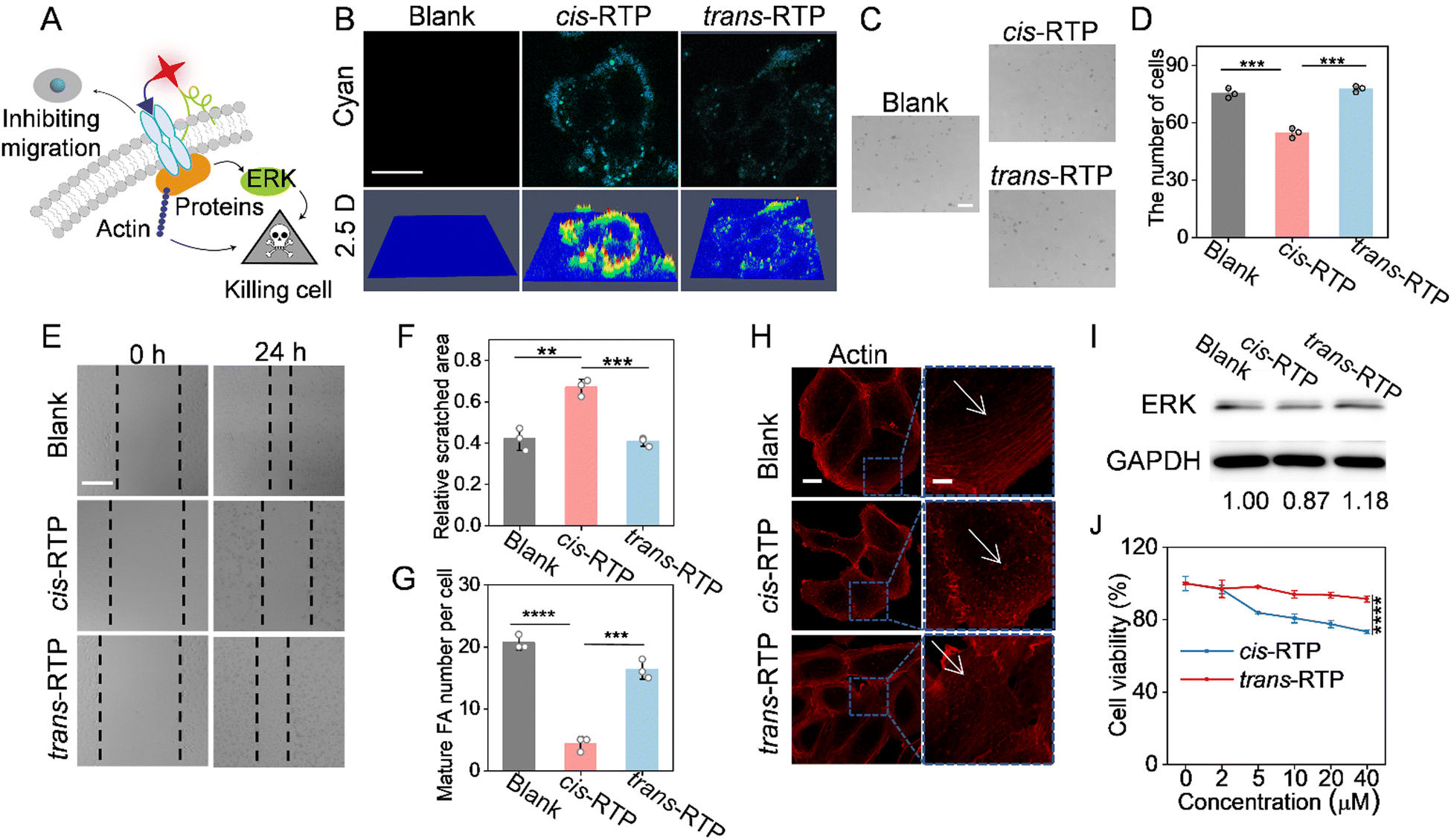 | ||
| Fig. 4 (A) Scheme of effects of probes on cell adhesion, migration and cell growth. (B) Confocal laser scanning microscopy (CLSM) images and 2.5D images of SKOV3 cells incubated with cis-RTP/trans-RTP (40 μM) for 1 h. Scale bar: 20 μm. (C) Adhesion result of suspended SKOV3 cells incubated with cis-RTP/trans-RTP (40 μM) for 1 h and then incubated with fresh medium for 4 h and (D) corresponding quantitative data of adherent SKOV3 cells in three separate fields. Scale bars: 200 μm. (E) Cell migration results of SKOV3 cells incubated with cis-RTP/trans-RTP (40 μM) for 1 h and then incubated with fresh medium and (F) closure degree of the scratched area was expressed as a percentage of the initial scratched area. Scale bar: 100 μm. (G) Quantification of dorsal bundle number and (H) CLSM images of the actin in SKOV3 cells incubated with cis-RTP/trans-RTP (40 μM) for 1 h and then stained with Actin-Tracker Red-555. The scale bar of actin images was 20 μm and scale bar of enlarged images was 5 μm. (I) Western blots of the expression levels of ERK in SKOV3 cells. SKOV3 cells were incubated with cis-RTP/trans-RTP (40 μM) for 1 h, and incubated with fresh medium for 24 h. (J) Cell viability of SKOV3 cells after incubation with cis-RTP/trans-RTP at different concentrations for 24 h. Data are expressed as mean ± SD; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. | ||
To further evaluate the effects of the cis/trans structures on intracellular pathways, an actin staining experiment was carried out (Fig. 4G, H and Fig. S23, ESI†). The results showed that the number of dorsal actin fibers in cells treated with different probes differed. Further quantitative results showed that the number of dorsal actin fibers per cell in the cis-RTP group was less than 10, which was much lower than that in the other groups. Simultaneously, we investigated the expression level of the kinase ERK involved in the regulation of extracellular signals using western blots (Fig. 4I). Similarly, the expression level of ERK in cis-RTP is lower than that in trans-RTP. These results reconfirmed that due to the higher affinity to integrin αvβ3 on the cell membrane, cis-RTP could have a more obvious influence on the intracellular pathways. Moreover, cell viability was measured by a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. After 24 h of treatment, the cell viability of cis-RTP at 40 μM was less than 80%, while the cell viability of trans-RTP was basically unaffected (Fig. 4J), illustrating that by affecting the intracellular actin and ERK pathways, cis-RTP could ultimately inhibit tumor cell growth to some extent, while trans-RTP was essentially non-toxic to the cells.
Conclusions
In conclusion, by distinguishing the cis/trans isomers, we fully use the contribution from membrane interactions for transmembrane protein recognition. Due to the difference in the spatial structure, cis-RTP could bind to the integrin αvβ3 on the cell membrane more strongly than trans-RTP, as evidenced by ITC and stimulation experiments. With tighter interaction with integrin αvβ3, cis-RTP was able to further influence the cell adhesion, migration, and intracellular pathway proteins. Simultaneously, cis-RTP exhibited toxicity to cells to some extent by affecting the intracellular actin and ERK pathways. These probes might be efficient fluorescent agents for membrane imaging, and the differentiation provided a promising strategy for modulating the interaction with proteins. This study also demonstrated that the difference between cis/trans isomers in biological applications could be an important reference for the development of more efficient and high-quality probes.Author contributions
J.-J. H., J. Yang and X. L. conceived the project and designed the experiments. F. X. and X. L. supervised the research. J. Yang and G. L. performed the experiments and analyzed the data. Y. L., Z. Z., F. X. and X. L. helped to analyze the data and provided valuable advice. J.-J. H., J. Yang and X. L. co-wrote the manuscript.Data availability
The data supporting this article have been included as part of the ESI.†Conflicts of interest
There are no conflicts to declare.Acknowledgements
This work was financially supported by the National Key R&D Program of China (2020YFA0211200, 2021YFA1200403), the National Natural Science Foundation of China (22090050, 21974128, 21874121), the Joint NSFC-ISF Research Grant Program (Grant No: 22161142020), Hubei Provincial Natural Science Foundation of China (2020CFA037). The authors would like to thank the high-performance computing platform of China University of Geosciences, Innovation Academy for Precision Measurement Science and Technology Chinese Academy of Sciences.Notes and references
- L. Jiang, M. Wang, S. Lin, R. Jian, X. Li, J. Chan, G. Dong, H. Fang, A. E. Robinson and M. P. Snyder, Cell, 2020, 183, 269 CrossRef PubMed.
- M. Labib, E. H. Sargent and S. O. Kelley, Chem. Rev., 2016, 116, 9001 CrossRef CAS PubMed.
- M. Labib and S. O. Kelley, Nat. Rev. Chem., 2020, 4, 143 CrossRef PubMed.
- J. A. Wells and C. L. McClendon, Nature, 2007, 450, 1001 CrossRef CAS PubMed.
- S. R. Tzeng and C. G. Kalodimos, Nature, 2012, 488, 236 CrossRef CAS.
- B. M. Kuenzi, L. L. R. Rix, P. A. Stewart, B. Fang, F. Kinose, A. T. Bryant, T. A. Boyle, J. M. Koomen, E. B. Haura and U. Rix, Nat. Chem. Biol., 2017, 13, 1222 CrossRef CAS PubMed.
- J. Zhou and J. Rossi, Nat. Rev. Drug Discovery, 2017, 16, 181 CrossRef CAS PubMed.
- K. Zhang, R. Deng, Y. Sun, L. Zhang and J. Li, Chem. Sci., 2017, 8, 7098 RSC.
- J. Bos and T. W. Muir, J. Am. Chem. Soc., 2018, 140, 4757 CrossRef CAS.
- H. Li, C. H. Lee and I. Shin, Chem. Commun., 2022, 58, 6336 RSC.
- Z. Pode, R. Peri-Noar, J. M. Georgeson, T. Ilani, V. Kiss, T. Unger, B. Markus, H. M. Barr, L. Motiei and D. Margulies, Nat. Nanotechnol., 2017, 12, 1161 CrossRef.
- A. M. Joffrin and L. C. Hsieh-Wilson, J. Am. Chem. Soc., 2020, 142, 13672 CrossRef PubMed.
- J. Tan, M. Zhao, J. Wang, Z. Li, L. Liang, L. Zhang, Q. Yuan and W. Tan, Angew. Chem., Int. Ed., 2019, 58, 1621 CrossRef PubMed.
- H. N. Chang, B. Y. Liu, Y. K. Qi, Y. Zhou, Y. P. Chen, K. M. Pan, W. W. Li, X. M. Zhou, W. W. Ma, C. Y. Fu, Y. M. Qi, L. Liu and Y. F. Gao, Angew. Chem., Int. Ed., 2015, 54, 11760 CrossRef PubMed.
- F. Xia, J. Wu, X. Wu, Q. Hu, J. Dai and X. Lou, Acc. Chem. Res., 2019, 52, 3064 CrossRef PubMed.
- J. Yang, J. Wei, F. Luo, J. Dai, J. J. Hu, X. Lou and F. Xia, Top. Curr. Chem., 2020, 378, 47 CrossRef PubMed.
- J. J. Hu, W. Jiang, L. Yuan, C. Duan, Q. Yuan, Z. Long, X. Lou and F. Xia, Aggregate, 2021, 2, 48 CrossRef.
- A. Loas and B. L. Pentelute, Chem. Rev., 2020, 120, 3049 CrossRef CAS PubMed.
- Z. Zhang, W. Zhu and T. Kodadek, Nat. Biotechnol., 2000, 18, 71 CrossRef CAS PubMed.
- T. A. Whitehead, Science, 2017, 358, 450 CrossRef CAS PubMed.
- J. Dai, J. J. Hu, X. Dong, B. Chen, X. Dong, R. Liu, F. Xia and X. Lou, Angew. Chem., Int. Ed., 2022, 61, e202117798 CrossRef CAS PubMed.
- J. Dai, X. Dong, R. Liu, B. Chen, X. Dong, Q. Wang, J. J. Hu, F. Xia and X. Lou, Biomaterials, 2022, 285, 121528 CrossRef CAS PubMed.
- C. Duan, J. J. Hu, R. Liu, J. Dai, M. Duan, L. Yuan, F. Xia and X. Lou, Angew. Chem., Int. Ed., 2021, 60, 18280 CrossRef CAS.
- B. Hu, N. Song, Y. Cao, M. Li, X. Liu, Z. Zhou, Q. Shi and Z. Yu, J. Am. Chem. Soc., 2021, 143, 13854 CrossRef CAS PubMed.
- S. Nishimua, K. Nishida and M. Tanaka, Chem. Commun., 2022, 58, 505 RSC.
- Y. Huang, M. Nawatha, I. Liveh, J. M. Rogers, H. Sun, S. K. Singh, A. Ciechanover, A. Brik and H. Suga, Chem. – Eur. J., 2020, 26, 8022 CrossRef CAS PubMed.
- M. A. Yıldırım, K. I. Goh, M. E. Cusick, A. L. Bara-bási and M. Vidal, Nat. Biotechnol., 2007, 25, 1119 CrossRef.
- I. Levental and E. Lyman, Nat. Rev. Mol. Cell Biol., 2023, 24, 107–122 CrossRef CAS.
- D. Cox, M. Brennan and N. Moran, Nat. Rev. Drug Discovery, 2010, 9, 804 CrossRef CAS PubMed.
- H. Hamidi and J. Ivaska, Nat. Rev. Cancer, 2018, 18, 533–548 CrossRef CAS PubMed.
- R. Nishiuchi, N. Sanzen, S. Nada, Y. Sumida, Y. Wada, M. Okada, J. Takagi, H. Hasegawa and K. Sekiguchi, Proc. Natl. Acad. Sci. U. S. A., 2005, 102, 1939 CrossRef.
- S. J. Shattil, C. Kim and M. H. Ginsberg, Nat. Rev. Mol. Cell Bio., 2010, 11, 288 CrossRef.
- Y. Cheng, C. Sun, R. Liu, J. Yang, J. Dai, T. Zhai, X. Lou and F. Xia, Angew. Chem., Int. Ed., 2019, 58, 5049 CrossRef PubMed.
- H. Li, Q. Yao, F. Xu, Y. Li, D. Kim, J. Chung, G. Baek, X. Wu, P. F. Hillman, E. Y. Lee, H. Ge, J. Fan, J. Wang, S. J. Nam, X. Peng and J. Yoon, Angew. Chem., Int. Ed., 2020, 59, 10186 CrossRef.
- H. B. Cheng, Y. Li, B. Z. Tang and J. Yoon, Chem. Soc. Rev., 2020, 49, 21 RSC.
- R. O. Hynes, Cell, 2002, 110(21), 673 CrossRef PubMed.
- J. D. Humphries, A. Byron and M. J. Humphries, J. Cell Sci., 2006, 119, 3901 CrossRef PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d4tb01801j |
| This journal is © The Royal Society of Chemistry 2024 |