
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceMaternal high-fat diet during lactation reduces sialylated milk oligosaccharides and shapes early-life microbiota in rat offspring†
Lucie Marousez‡
a,
Marko Verce‡ bc,
Laure Dubernata,
Thameur Rakzade,
Patrice D. Canibcf,
Frédéric Gottrandag,
Delphine Leyag,
Amandine Everardbc,
Delphine Eberlé§
h and
Jean Lesage§*a
bc,
Laure Dubernata,
Thameur Rakzade,
Patrice D. Canibcf,
Frédéric Gottrandag,
Delphine Leyag,
Amandine Everardbc,
Delphine Eberlé§
h and
Jean Lesage§*a
aUniversity of Lille, Inserm, CHU Lille, U1286-INFINITE-Institute for Translational Research in Inflammation, F-59000 Lille, France. E-mail: jean.lesage@univ-lille.fr
bMetabolism and Nutrition Research Group, Louvain Drug Research Institute (LDRI), UCLouvain, Université catholique de Louvain, Brussels, Belgium
cWalloon Excellence in BIOtechnology (WELBIO) Department, WEL Research Institute (WELRI), Wavre, Belgium
dUniversity of Lille, ULR 2694 METRICS, F-59000 Lille, France
eCHU Lille, Department of Pediatrics, Lille University Hospital, F-59000 Lille, France
fInstitute of Experimental and Clinical Research (IREC), UCLouvain, Université catholique de Louvain, 1200 Brussels, Belgium
gDivision of Gastroenterology Hepatology and Nutrition, Department of Paediatrics, Jeanne de Flandre Children's Hospital, CHU Lille, F-59000 Lille, France
hUniversity of Lille, Inserm, CHU Lille, Institut Pasteur de Lille, U1011-EGID, F-59000, Lille, France
First published on 5th June 2025
Abstract
Human milk oligosaccharides (HMOs) promote the development of the infant gut microbiota, notably through the growth of beneficial bacteria. However, the interaction between maternal diet, HMOs and the offspring's gut microbiota remains incompletely understood. We hypothesize that a high-fat (HF) diet influences the composition of HMOs and shapes gut microbiota in offspring. In lactating rat mothers fed with a HF or a low-fat (LF) diet, we measured at mid-lactation, the levels of milk sialylated oligosaccharides (SL-MOs) and the expression of enzymes for synthesizing SL-MOs in the mammary gland. In rat pups from LF or HF mothers, we analysed the caecal microbiota and measured levels of faecal short-chain fatty acids (SCFAs). In addition, in humans, we quantified 24 HMOs in the colostrum of lean and obese mothers. In rat mothers, HF feeding significantly reduced the breast milk 6′SL level but not the 3′SL one and increased the gene-expression level of the St3Gal1 enzyme in the mammary gland. In HF rat pups, the caecal microbiota was drastically modified with increases in α- and β-diversities as well as in the abundance of Parabacteroides and Lachnospiraceae. Levels of faecal SCFAs including acetate, butyrate, isobutyrate, valerate and isovalerate were also increased in HF pups. In humans, maternal obesity only slightly increased DFLNHa concentration in the colostrum. In conclusion, our data suggest that a hyperlipidic diet during lactation may reduce the level of sialylated oligosaccharides in breast milk and alter the abundance of several groups of bacteria in the infant's gut. We also demonstrated that, in humans, obesity did not highly affect levels of HMOs in the colostrum reinforcing previous findings suggesting that being overweight does not influence drastically the HMO profile in breast milk.
1. Introduction
To establish advanced strategies for supporting infant health, a deeper understanding of the mechanisms implicated in early-life intestinal microbial colonization and development is essential. Indeed, recent evidence has increasingly highlighted that the development of the gut microbiota during infancy is a pivotal process with profound implications for both short- and long-term health.1–3 In infants, the early gut microbiota has been shown to play numerous important roles, including aiding in nutrient metabolism, producing short-chain fatty acids (SCFAs), supporting the development of the gut barrier and immune system, as well as protecting against pathogens.2,3 Breastfeeding is widely recognized for its pivotal role in shaping the early development of the infant gut microbiota, a process influenced by specific bioactive components from human milk.4,5 Among these components, human milk oligosaccharides (HMOs) have gained remarkable interest in recent years due to their diverse biological functions, particularly their ability to promote the growth of beneficial bacteria such as Bifidobacterium species in the infant gut.6,7HMOs consist of a combination of five monosaccharides including glucose, galactose, N-acetylglucosamine, fucose, and sialic acid.6,7 These monosaccharides are combined in various ways to form different core HMOs that can be further modified by sialic acid and fucose to generate sialylated and/or fucosylated HMOs.6,7 HMOs are divided into several groups, i.e., fucosylated (neutral) and non-fucosylated (neutral) oligosaccharides and sialylated (acidic) molecules.8 Some minor HMOs also contain both fucose and sialic acid molecules, forming an additional subgroup.9 The highest concentration of HMOs is found in colostrum, up to 20 g L−1. Thereafter, as the composition of breast milk changes during lactation, the concentration of HMOs decreases to 7–15 g L−1 in mature breast milk.6,7 Human milk has a complex composition of HMOs with around 150 different molecules identified so far. However, fewer than 50 of these HMOs are present in significant amounts and proposed as exerting significant biological roles.8
In breastfed infants, the majority of HMOs (close to 99%) reach the large intestine due to their resistance to acidic pH and their inability to be hydrolyzed by pancreatic and intestinal enzymes.7 In the colon, these molecules serve as substrates for bacterial metabolism by specific gut microbiota members, notably from the genus Bifidobacterium.6 However, some Bacteroides and Akkermansia species are also utilizers of HMOs.7,10 Among the early gut colonizers that specifically ferment HMOs, several species of Bifidobacterium, such as B. bifidum, B. breve, and B. longum subsp. infantis used HMOs as their sole carbon source.11 For the genus Bacteroides, B. fragilis, B. vulgatus, B. caccae, and B. thetaiotaomicron are also important utilizers of HMOs.12 Furthermore, other bacterial species, such as Lactobacillus species, also benefit from HMOs for their development.13 HMOs also act directly on intestinal and immune cells and exert antiadhesive and antimicrobial effects, as well as beneficial immunomodulatory roles.7,14
The composition and content of HMOs in human milk can vary significantly due to various factors, such as maternal genetics and health, the stage of lactation, but also environmental factors, such as diet, season and geographical location.7,15,16 However, there is a limited number of experimental and clinical studies available, particularly those focusing on the effect of maternal diet on HMOs. A deeper understanding of the functional interactions between specific nutrients of maternal diet and HMOs is crucial for developing future dietary interventions in lactating women to promote infant health. To address this, we established a rat model of maternal high-fat (HF) diet administered exclusively during lactation. We measured levels of sialylated milk oligosaccharides (SL-MOs), which are the main oligosaccharides in rat milk,7 and analyzed the gene expression of several enzymes for synthesizing sialylated oligosaccharides in the mammary glands of lactating rats. Additionally, to investigate the putative impact of changes in SL-MOs on pup gut microbiota, we investigated the caecal microbiota composition in rat pups at mid-lactation as well as levels of faecal short-chain fatty acids (SCFAs). Finally, to assess the translational application of our results using a clinical cohort, we measured the level of 24 HMOs in the colostrum of mothers with or without obesity.
2. Materials and methods
2.1. Animal model and samples
Lactating female Wistar rats (Charles River Laboratories, L'Arbresle, France) suckled 8 pups each (4 males and 4 females), and received either a HF diet (n = 7 litters; 60% kcal from fat, D12492, research diet, New Brunswick, USA) or a control low-fat (LF) diet (n = 7 litters; 10% kcal from fat, D12450J, Research Diet) exclusively during the lactation period (from postnatal day 1 (P1) to P12) as previously described.17 The entire composition of HF/LF diets is reported in ESI Table S1.† Maternal and pup body weights were measured from P1 to P12 of lactation. Breast milk (BM) was collected from anesthetized P12 mothers after oxytocin administration as previously described.17 Briefly, mothers were anesthetized at P12 on isoflurane and mammary glands (MG) were externally gently massaged for BM collection after oxytocin administration intraperitoneally (2U, Sigma-Aldrich, St. Louis, USA). For MG collection, mothers were euthanized after BM collection by intracardiac blood puncture performed on anesthetized animals. MGs were collected post-mortem, flash frozen in liquid nitrogen and stored appropriately at −80 °C for RNA analysis as previously described.17At P12, only male pups of both LF and HF groups (n = 22–28 per group) were euthanized for sample collection as previously described17 in line with a previous study of our group focusing on the programming of obesity only in male pups using this model. Caecal and faecal contents were collected and immediately flash-frozen and stored at −80 °C. Animal protocols were conducted in accordance with the principles of laboratory animal care (European Communities Council Directive of 1986, 86/609/EEC) and approval was received from the “Région Hauts-de-France” ethical committee for animal experimentation (Comité d’éthique en expérimentation animale du Nord - Pas de Calais; no. CEEA-075).
2.2. Human cohort and samples
To investigate, in humans, whether a high amount of adipose tissue in lactating mothers can affect breast milk HMOs, a cohort of breastfeeding mothers composed of mothers with or without obesity (BMI > 30 or BMI between 18 and 25, respectively) (n = 15 per group) was used (ESI Table S2†) at the Jeanne de Flandre Maternity Hospital (Lille, France). Inclusion criteria were women between 18 and 42 years of age, with social insurance, presenting a single pregnancy with a term between 35 and 40 SA and consenting after informed consent. Exclusion criteria were: pre-pregnancy diabetes, gestational diabetes, hypertension, pre-eclampsia, IUGR <10th percentile, no medication and long-term medication, drug use, renal insufficiency, hepatic or cardiac insufficiency, a history of bariatric surgery, foetal anoxia with an umbilical cord pH below 7.0, a prenatally diagnosed genetic or chromosomal abnormality of the mother and/or newborn, or a prenatally diagnosed foetal malformation. In this small clinical cohort only colostrum samples were collected 2–3 days after delivery in the morning (≈11 am) without any fasting. In this cohort, mature breast milk was not available making comparison to ‘mature’ P12 rat milk not possible. All samples were frozen at −80 °C until further analysis. Patient's informed consent was obtained at the beginning of the study. Clinical protocols were approved by the “Nord Ouest IV” committee for the protection of persons and received the approval number 2015 A01696 43.2.3. Quantification of oligosaccharides in rat and human milk
We quantified the level of two SL-MOs in P12 rat milk (3′-sialyllactose (3′SL) and 6′-sialyllactose (6′SL)) and the level of 24 HMOs in human milk from our clinical cohort using a previously described and validated method.18 All milk samples were shipped on dry ice to Neotron S.p.A. (Italy) for oligosaccharides quantification by liquid chromatography with fluorescence detection after labelling with 2-aminobenzamide (2AB) as described by Austin & Benet (2018).18 An Ultimate 3000-RS ultra-high performance liquid chromatography (UHPLC) system equipped with an RF-2000 fluorimeter (FLD) and a 2-way 10-port high pressure switching valve (all from Thermo Fisher Scientific, Waltham, USA) was used. The columns were an Acquity BEH Glycan (1.7 μm, 2.1 × 150 mm) and VanGuard BEH amide (1.7 μm, 2.1 × 50 mm) both from Waters Corporation (Milford, USA). The guard column was installed between the injector and the 10-port valve. Eluent A was acetonitrile and eluent B was ammonium formate (50 mmol L−1). After injection, the sample was directed to the guard column with flow diverted to the waste using an eluent composition of 5% B for 2.5 min at a flow rate of 0.5 mL min−1. Then the flow was directed through the analytical column for elution of oligosaccharides with the following gradient: 2.4 min with 10% B (isocratic), then ramp % B from 10% to 18% in 27.2 min then 18.0–19.5% in 16.0 min, 19.5%–22.0% in 13.4 min and 22.0%–25.4% in 27.5 min. The flow rate was 0.5 mL min−1 and the analytical column was kept at 55 °C. Detection was performed using an excitation wavelength of 330 nm and emission wavelength of 420 nm. Ten HMOs were quantified using genuine standards of HMOs with known purity assessed using quantitative nuclear magnetic resonance spectroscopy (Elicityl, France), namely 2′-fucosyllactose (2′FL), 3-fucosyllactose (3FL), lacto-N-tetraose (LNT), lacto-N-neotetraose (LNnT), 3′-sialyllactose (3′SL), 6′-sialyllactose (6′SL), lacto-N-fucopentaose-I (LNFP-I), lacto-N-fucopentaose-V (LNFP-V), and lacto-N-neofucopentaose (LNnFP). The remaining HMOs were quantified against maltotriose of known purity (Sigma-Aldrich, Germany), assuming equimolar response factors. The full names and abbreviations of all quantified rat SL-MOs and HMOs are indicated in the legend of Fig. 2 and 5.2.4. Real-time quantitative polymerase chain reaction (RT-qPCR)
Total ribonucleic acid (RNA) was extracted from P12 rat mammary glands (n = 7 per group) using RNeasy Mini Kits (Qiagen, Hilden, Germany). Reverse transcription was performed using an iScript kit (Bio-Rad Laboratories, Hercules, USA). Using specific primers (ESI Table S3†), mRNA levels of three sialyltransferases implicated in the synthesis of milk 3′SL and 6′SL were quantified with SYBR Green (Thermo Fisher Scientific, Illkrich-Graffenstaden, France) in a StepOne system (Thermo Fisher Scientific). The expression of transcripts was analyzed by the 2−ΔCt method using the mean of cyclophilin B as reference genes as previously described.172.5. Caecal microbiota analysis in P12 rat pups
DNA was extracted from male rat pups caecal samples (n = 12 per group) using a QIAamp DNA Stool Mini Kit (Qiagen) according to the manufacturer's instructions with modifications,19 followed by the amplification of the 16SrRNA gene V1–V3 regions using the primers 27Fmod (5′-AGRGTTTGATCMTGGCTCAG-3′) and 519Rmodbio (5′-GTNTTACNGCGGCKGCTG-3′). Amplicon sequencing was performed using an Illumina MiSeq platform at MRDNA (Shallowater, TX, USA). QIIME2 (q2cli 2021.4.0)20 was used for sample processing, starting with Cutadapt21 for primer removal, using the following parameters: overlap = 5, error rate = 0.1, discard untrimmed reads. DADA222 was used for denoising of pre-merged reads using the default parameters in QIIME2 with the following modifications: maximum expected errors = 2, truncation length = 453 nt. A classifier for the V1–V3 region was built using the SILVA 138.1 SSURef NR99 database,23 using RESCRIPt24 for dereplication and trimming accordingly. Taxonomy assignment and mapping to the rat genome with BLAST in QIIME2 were used to detect off-target ASVs, resulting in the removal of 14 ASVs. The dataset consisted of 354 ASVs in 24 samples, with between 45![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 582 and 67263 sequence reads per sample, averaging at 56452. The R package collection “tidyverse” was used for graphics visualisation.25 The sequencing data were submitted to the European Nucleotide Archive (ENA/EBI) and are available under the study accession number PRJEB83305. Quantification of total bacteria was carried out by qPCR with universal bacterial primers (338F: ACTCCTACGGGAGGCAGCAG, 518R: ATTACCGCGGCTGCTGG), with the StepOnePlus real-time PCR system and software (Applied Biosystems, Den Ijssel, The Netherlands) using GoTaq qPCR SYBR green mix (Promega, Madison, Wisconsin, USA), according to the manufacturer's instructions. All samples were run in duplicate in a single 96-well reaction plate. The cycle threshold of each sample was compared with a standard curve made by serially diluting genomic DNA isolated from a pure culture of the type strain of Lactobacillus acidophilus (DSM 20079 01-21) (BCCM/LMG, Ghent, Belgium; DSMZ, Braunshweig, Germany). The absolute abundances of individual bacterial genera were estimated by multiplying their relative abundance by total bacterial density as described previously.26
582 and 67263 sequence reads per sample, averaging at 56452. The R package collection “tidyverse” was used for graphics visualisation.25 The sequencing data were submitted to the European Nucleotide Archive (ENA/EBI) and are available under the study accession number PRJEB83305. Quantification of total bacteria was carried out by qPCR with universal bacterial primers (338F: ACTCCTACGGGAGGCAGCAG, 518R: ATTACCGCGGCTGCTGG), with the StepOnePlus real-time PCR system and software (Applied Biosystems, Den Ijssel, The Netherlands) using GoTaq qPCR SYBR green mix (Promega, Madison, Wisconsin, USA), according to the manufacturer's instructions. All samples were run in duplicate in a single 96-well reaction plate. The cycle threshold of each sample was compared with a standard curve made by serially diluting genomic DNA isolated from a pure culture of the type strain of Lactobacillus acidophilus (DSM 20079 01-21) (BCCM/LMG, Ghent, Belgium; DSMZ, Braunshweig, Germany). The absolute abundances of individual bacterial genera were estimated by multiplying their relative abundance by total bacterial density as described previously.26
2.6. Levels of faecal SCFAs in P12 rat pups
The faecal content of male LF and HF pups at P12 (n = 10–12 per group) was homogenized in 1.5 mL of 0.005 M NaOH solution, including internal standards (acetate-D3, propionate-D2, butyrate-13C2 and valerate-D9) using Precellys equipment. Total DNA was extracted following several steps and SCFAs were measured by chromatography-mass spectrometry (GC/MS) analysis as previously described.27 The GC/MS analysis comprised a phase of liquid injection at 260 °C in the split mode and a separation phase on a 50 m × 0.25 mm, 0.25 μm, DB-5 ms capillary column. Quantification was performed with a single quadrupole using electron impact ionization.2.7. Statistical analysis
Statistical analysis (indicated in the figure legends) was performed with GraphPad Prism 9.0 software. Grubb's test was used to detect any outliers. Normality of variables was evaluated by a D'Agostino–Pearson test. If needed, a log10 transformation of data was performed to reach normality. Statistical differences were tested by a Mann–Whitney or Student's t-test according to sample normality assessment results. The gut microbial data were processed with QIIME2. According to the rarefaction curves, the minimum numbers of sequences per sample represented a sufficient sampling depth. Principal coordinate analyses (PCoA) were performed on weighted and unweighted UniFrac distances (wUniFrac, unwUniFrac28). Differences between group α-diversity metrics were assessed with the Kruskal–Wallis test. PERMANOVA and PERMDISP, both at 9999 permutations, were used for statistical evaluations of beta-diversity. Differentially abundant ASVs and genera were detected using ANCOM-BC.29 To obtain a Spearman correlation matrix between the concentrations of milk 3′SL and 6′SL and abundances of ASVs or taxa in the pup gut microbiota, the relative frequencies of ASVs and taxa were transformed to the absolute scale using the 16S rRNA gene-based quantitative PCR measurements of the total bacterial load. These data were combined for the creation of an ASV- or taxon-level correlation matrix using the R package Hmisc and the rcorr function. A Benjamini–Hochberg correction was used to correct the resulting p-values, whereby the corrected value of 0.05 was considered as the significance threshold.
3. Results
3.1. Maternal HF diet increases the body weight of P12 rat pups and reduces the breast milk levels of 6′SL
At P12, the body weight (BW) of HF and LF mothers was similar (Fig. 1A). However, male HF pups displayed 7% higher BW than LF pups (Fig. 1B). The concentrations of the two major rat SL-MOs (3′SL and 6′SL) were analysed at P12. Maternal HF diet did not significantly affect 3′SL levels but there was a trend towards a slight reduction of this compound (p = 0.202, Fig. 2A and ESI Table S4†). However, in milk of HF mothers, 6′SL concentrations were reduced by 15% (p = 0.007, Fig. 2A and ESI Table S4†). The gene-expression levels of three enzymes involved in 3′SL and 6′SL syntheses pathways were measured in the MGs of mothers at P12. MG St3Gal1 mRNA expression was significantly increased in HF mothers (p = 0.039, Fig. 2B) whereas St3Gal4 and St6Gal1 mRNA expressions were similar between LF and HF mothers (p = 0.872, p = 0.221 respectively). | ||
| Fig. 1 Effects of HF diet during lactation on maternal body weight (A) and body weight of rat pups (B) at postnatal day 12 (P12) (n = 7 per group (A) and n = 22–28 per group (B)). Mean ± SEM. ***p < 0.001 HF vs. LF group by the Mann–Whitney test. | ||
 | ||
| Fig. 2 Breast milk levels of 3′SL and 6′SL in P12 lactating rat mothers (A) and gene-expression levels of the three enzymes St3Gal1, St3Gal4 and St6Gal1, which are involved in their syntheses in the mammary glands of LF and HF mothers at P12 (B) (n = 7 per group). Mean ± SEM. *p < 0.05, **p < 0.01 HF vs. LF group by the Mann–Whitney test; 3′/6′SL is 3′/6′-sialyllactose. | ||
3.2. Maternal HF diet alters the caecal microbiota of P12 pups and increases their content of faecal SCFAs
The gut microbiota of P12 rat pups exhibited limited diversity and was mainly composed of Proteobacteria (mainly Escherichia–Shigella) and Firmicutes (mainly Ligilactobacillus). Bacteroidota were largely represented only in the caecal content of HF pups (Fig. 3A). The enhanced presence of Bacteroidota was reflected in significantly higher α-diversity metrics linked to richness and phylogenetic diversity in HF pups (Fig. 3B–E). The difference in gut microbiota composition was also significant between the two groups in terms of β-diversity, regardless of whether or not ASV relative frequencies were taken into account (PERMANOVA, p = 0.0001 for wUniFrac and unwUniFrac; Fig. 3F and G). The dispersion within the HF group was significantly higher than in the LF group when relative frequencies were taken into account (PERMDISP, wUniFrac, p = 0.0014). Differential abundance analysis with ANCOM-BC revealed 23 significant genera: Parabacteroides, unknown Lachnospiraceae, Negativibacillus, UBA1819, unknown Ruminococcaceae, Faecalitalea, Butyricicoccus, Romboutsia, Colidextribacter, [Eubacterium] nodatum group, Hungatella, unknown Clostridia, and Frisingicoccus were more abundant in HF pups, whereas Enterococcus, Terrisporobacter, Marvinbryantia, Clostridium sensu stricto 1, Escherichia–Shigella, Anaerotruncus, Lachnoclostridium, Lactobacillus, unknown Lactobacillaceae, and the [Eubacterium] coprostanoligenes group were less abundant in this group (Fig. 3A). Moreover, several ASVs and taxa within the pup gut microbiota significantly correlated with the concentrations of 3′SL and 6′SL in maternal breast milk (ESI Fig. S1A and B†). In particular, twelve significant correlations between ASVs and SL-MOs were identified (ESI Table S4†). Two ASVs belonging to Clostridium sensu stricto 13 and Escherichia–Shigella correlated positively with 3′SL, while two ASVs belonging to unknown or uncultured Ruminococcaceae correlated negatively with both 3′SL and 6′SL levels. Three ASVs belonging to Enterococcus and two ASVs belonging to Blautia and Clostridium sensu stricto 1 correlated positively with 6′SL; an additional ASV belonging to Parabacteroides correlated negatively with 6′SL. Four significant taxon-level correlations with the two SL-MOs were found: unknown Ruminococcaceae correlated negatively with both 3′SL and 6′SL, whereas Enterococcus and Anaerotruncus correlated positively with 6′SL. Additionally, amounts of five major SCFAs including acetate, butyrate, isobutyrate, valerate and isovalerate were increased in the faeces of P12 HF pups (Fig. 4A–C, E and F). Although, faecal propionate was not significantly affected, a tendency for a similar increase in HF pups was observed (Fig. 4D). | ||
| Fig. 3 Analysis of the caecal microbiota of P12 rat pups fed by LF or HF mothers. (A) A genus-level taxonomy bar-plot for each sample in the LF and HF groups. ASVs assigned only to a taxonomic rank higher than genus were grouped and depicted at that rank. Taxa more or less abundant in the HF group compared to the LF group according to ANCOM-BC are framed red or blue in the legend, respectively. “Minorities” – taxa represented at <1.0% relative frequency in all samples. (B–E) Alpha-diversity metrics (Shannon index, Pielou's evenness, observed ASVs, Faith's phylogenetic diversity) with the Kruskal–Wallis p-values. (F and G) Principal coordinate analysis plots based on unweighted (F) and weighted (G) UniFrac distances, with PERMANOVA and PERMDISP p-values (n = 12 per group). | ||
 | ||
| Fig. 4 Levels of faecal SCFAs in P12 rat pups from LF and HF mothers. (A) Acetate, (B) butyrate, (C) isobutyrate, (D) propionate, (E) valerate and (F) isovalerate concentrations in faecal samples (n = 22–28 per group). Mean ± SEM. *p < 0.05; **p < 0.01; ***p < 0.001 HF vs. LF group by the Mann–Whitney test or Student's t-test. | ||
3.3. Levels of HMOs in colostrum are not affected by maternal obesity
We measured the levels of the 24 HMOs in our clinical cohort. Using data reported in Fig. 5 and ESI Table S5,† we were able to determine the secretor status of lactating mothers. Indeed, as high levels of both 2′FL and LNFP-I were found in all samples, in accordance with Tonon et al.,30 the presence of these specific HMOs at high levels is sufficient to characterize the maternal phenotype as secretor. Regarding the influence of maternal obesity, we did not observe major impacts of this pathology on the amount of these HMOs in colostrum. Only DFLNHa, which is present at a very low level in human milk of secretor mothers, was significantly increased by maternal obesity (Fig. 5A and ESI Table S5†). Similarly, a tendency of an increased level of DSLNT (P = 0.058) was observed in mothers with obesity (Fig. 5A and ESI Table S5†). The calculation of total levels of fucosylated, acidic and neutral non-fucosylated HMOs in milk samples did not show any significant modulation by obesity (Fig. 5B and ESI Table S5†).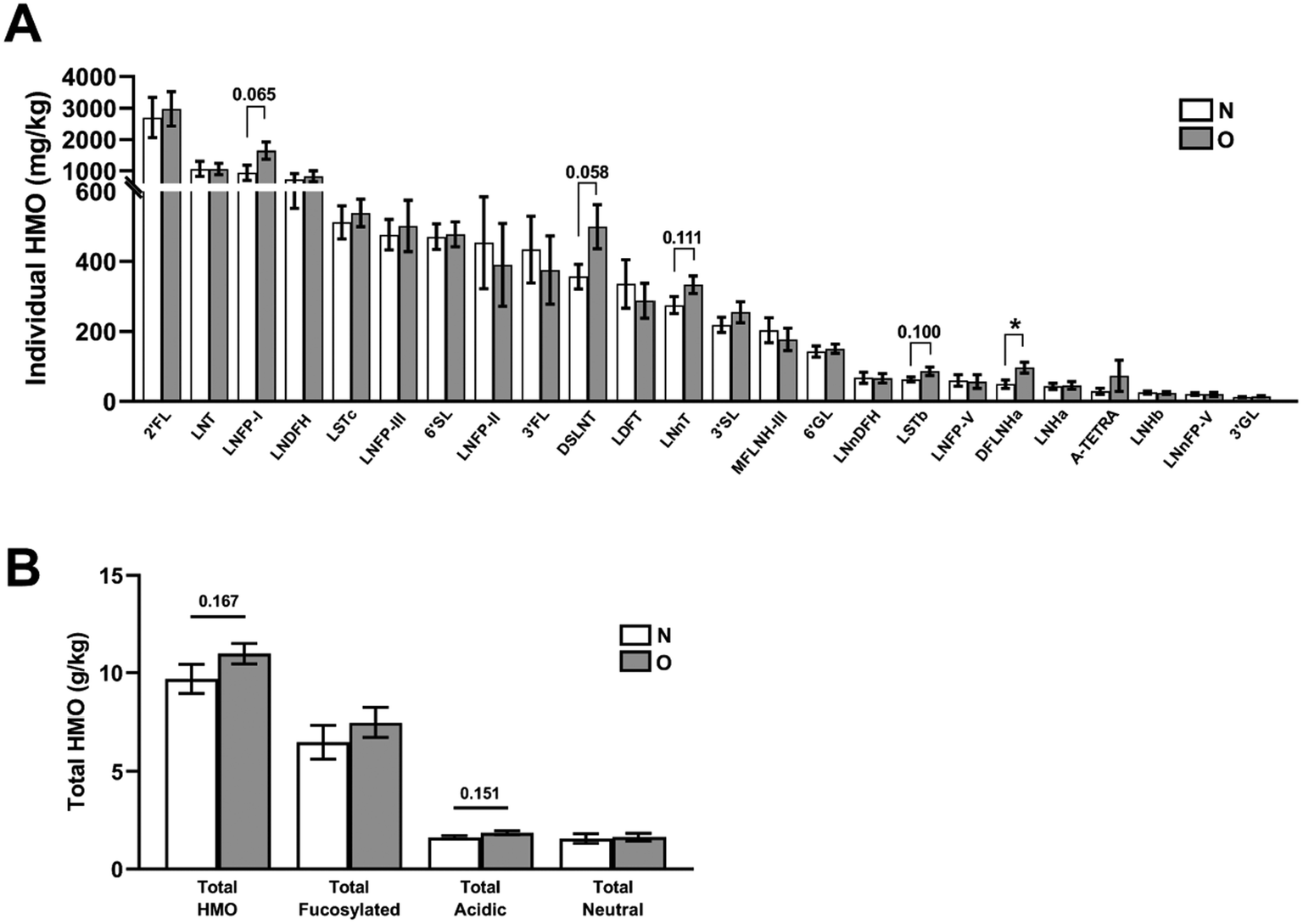 | ||
| Fig. 5 Individual and total concentrations of HMOs in the colostrum of a clinical cohort including normal-weight mothers (N) and mothers with obesity (O). Individual (A), total HMOs and types of HMOs (B) were quantified in colostrum samples. Total HMOs were calculated as the sum of all HMOs. The total fucosylated, acidic and neutral non-fucosylated HMOs were calculated by the sum of the HMOs belonging to a group and represented by g kg−1. A-TETRA, A-tetrasaccharide; DFLNHa, difucosylated lacto-N-hexaose a; DSLNT, disialyllacto-N-tetraose; 2′/3′FL, 2′/3′-fucosyllactose; 3′/6′GL, 3′/6′-galactosyloligosaccharide; LDFT, lactodifucotetraose (difucosyllactose); LNDFH, lacto-N-difucohexaose; LNFP-I/II/III/V, lacto-N-fucopentaose I/II/III/V; LNH a/b, lacto-N-hexaose a/b; LNnDFH, lacto-N-neodifucohexaose; LNnDFH, lacto-N-neodifucohexaose; LNnFP-V, lacto-N-neofucopentaose-V; LNnT, lacto-N-neotetraose; LNT, lacto-N-tetraose; LST b/c, sialyllacto-N-tetraose b/c; MFLNH-III, monofucosyllacto-N-hexose-III; 3′/6′SL, 3′/6′-sialyllactose; NF, neutral fucosylated; NnF, neutral non-fucosylated; S, sialylated (acidic). Mean ± SEM (n = 15 per group). *p < 0.05 O vs. N group by the Mann–Whitney test or Student's t-test. | ||
4. Discussion
The present study on a rat model of postnatal nutritional intervention demonstrated that HF feeding exclusively during lactation is sufficient to reduce levels of sialylated breast milk oligosaccharides and to shape the microbiota of rat offspring during the mid-lactation period. However, as only male offspring were investigated, this constitutes a major limitation of the present study. We further showed that maternal HF feeding during this brief period increased the body weight of rat offspring. A previous study using this experimental model revealed that the maternal HF diet drastically altered the fatty acid (FA) composition of breast milk resulting in a reduced proportion of medium-chain FAs, an increased proportion of long-chain FAs, elevated mono- and poly-unsaturated FAs and increased the n-6/n-3 ratio.17 In addition, this model was shown to program depot-specific adiposity in offspring, partly by modifying visceral white adipose tissue cellularity and the epigenetic landscape.17 This programming of adipose tissue development by maternal HF diet and the changes in breast milk lipids may explain the increased body weight observed in P12 rat pups in the present study. These hypotheses are in accordance with other previous studies that have demonstrated, for example, that the n-6/n-3 PUFA ratio in human milk is positively associated with early infant WAT deposition.31 Additionally, they are consistent with the presumed obesogenic role of n-6 poly-unsaturated FAs during the perinatal period as suggested by others authors.32That said, FAs are not the only milk components that could mediate the increase in body weight in the offspring. Indeed, breast milk oligosaccharides could also be involved in such effects knowing their effects on gut microbiota composition and the important roles of gut microbes in the regulation of host metabolism and body weight regulation in pups.1–3,19 Further studies are needed to assess whether specific microbes (e.g. Akkermansia muciniphila) are also implicated in the increased body weight of HF pups. The second key finding of this study is that a HF diet in lactating rat dams from postnatal day 1 (P1) to P12 is sufficient to reduce significantly the breast milk 6′SL concentration by 15%. In addition, the levels of 3′SL (the major SL-MOs measured in rat milk in this study) were slightly reduced in HF mothers, but this change was not statistically significant. Very few studies have investigated levels of milk oligosaccharides in rodents. Four studies have shown that 6′SL and at a lower level 3′SL are the two dominant oligosaccharides in mouse milk while very low levels or trace amounts of up to 19 other oligosaccharides have been detected in some other studies using mass spectrometry.33–36 Our findings in rat milk are aligned with results from Li et al.,33 showing that in contrast to mouse milk, 3′SL is the dominant milk oligosaccharide (concentration close to 7800 mg kg−1) and 6′SL is the second major milk oligosaccharide, which is far less abundant (concentration close to 36 mg kg−1). While we observed a reduction in milk 6′SL level in HF dams, we did not find any significant modulation of the gene-expression level of the sialyltransferase (St6Gal1) implicated in the synthesis pathway of 6′SL. Surprisingly, we observed an increased expression of the St3Gal1 enzyme in HF mothers without modulation of the St3Gal4 enzyme. This suggests that HF feeding may affect the metabolic pathway and/or the enzymatic activities implicated in the synthesis of milk containing sialylated oligosaccharides, as well as potentially other factors of the milk glycobiome knowing that sialyltransferases are also implicated in the synthesis of others milk glycoconjugate compounds.37 Consistent with our findings, Liu et al.36 observed that exposure to HF during eleven weeks including the gestational and lactation periods altered especially sialylated milk oligosaccharides in mice. Similarly, another study in humans found that a HF diet was similarly linked to a decrease in sialylated HMOs.38 Thus, these findings altogether support the concept that sialylated oligosaccharides in breast milk, especially 3′SL and 6′SL, may be reduced by a maternal hyperlipidic diet especially if this regimen takes place during the critical lactation period.
The third significant finding of this study is that maternal HF feeding exclusively during lactation alters the gut microbiota of P12 rat pups and increases the production of gut SCFAs. Despite the well-established beneficial roles of SCFAs (e.g., epithelial gut barrier function, anti-inflammatory effects, colonic homeostasis, appetite regulation),1–3 we found increased levels of SCFAs in the offspring of HF-fed dams. One potential explanation is that maternal HF diet exposure may have altered the gut microbiota of the offspring in a way that favours expansion of SCFA-producing taxa that could represent a compensatory microbial adaptation to an altered intestinal environment or nutrient landscape. Alternatively, the increased SCFAs might not necessarily confer beneficial effects in this context. The response of the host to SCFAs may be altered due to developmental programming by the maternal diet, potentially leading to impaired signalling or utilization of SCFAs, thereby suggesting lower absorption and/or utilization of SCFAs by the intestinal epithelial cells, eventually leading to higher concentrations of SCFAs in the faeces. These findings highlight the complex interplay between maternal diet, microbial composition, and host-microbiome interactions in early life. Future studies are needed to assess whether the observed increase in SCFAs exerts protective, detrimental, or neutral effects on offspring health outcomes. Concerning the gut microbiota of rat pups, our results demonstrate that a maternal HF diet largely influences the composition and activity (i.e., production of SCFAs) of the offspring gut microbiota with an increase in α-diversity and β-diversities as well as a differential abundance of several groups of bacteria, compared to the microbiota of LF diet offspring. Among the most affected bacterial genera, a high relative abundance of Parabacteroides only in HF offspring was found, which could be consistent with the increase in the production of SCFAs, in particular acetate, isovalerate, and propionate.39 These data confirmed recent findings showing that a maternal HF diet during lactation increases the α-diversity in the caecal gut microbiota of rat pups.40 Interestingly, it was also demonstrated in rats that it is the postnatal maternal diet rather than prenatal diet that predominantly shaped the offspring's gut microbiota.40 In addition, these authors also identified Parabacteroides as a characteristic feature of offspring gut microbiota from HF mothers.40 In our study, the increased abundance of one of the two main ASVs of Parabacteroides coincided with a lower concentration of 6′SL in rat milk. Besides Parabacteroides, other bacteria were also affected in our study, including those of Lachnospiraceae that were observed to increase in the gut microbiota of HF pups. Members of this diverse family of bacteria could also contribute to the increased pool of SCFAs, as many of them are butyrate producers.41 Interestingly, in a human cohort of breastfed infants, it was shown that both Parabacteroides and Lachnospiraceae were strongly and negatively correlated with faecal concentration of HMOs, reinforcing the well-described link that exists between breast milk oligosaccharides and gut microbiota composition in offspring.42
Finally, as previous findings in rats suggested that altered lipid status may reduce the contents of oligosaccharides in milk, we analysed the levels of 24 HMOs in the colostrum of mothers with or without obesity. Mature milk was not available in our clinical cohort as well as antibiotic treatment history in the last 3–6 months, which is a limitation of the present study to compare human results with previous findings in rats. Using this cohort, we did not find major modulations of concentrations of HMOs in the colostrum of mothers with a secretor phenotype and affected by obesity. However, we noted that the level of DFLNHa, which is a minor HMO in secretor-type mothers, was increased by maternal obesity. Our data are in accordance with those of others in clinical studies that have demonstrated that obesity did not affect drastically the levels of HMOs in transitional and mature breast milk.43–46
The findings from the present study may shed light on the consequences of a hyperlipidic diet during lactation on the contents of oligosaccharides in breast milk and on the gut microbiota in offspring. Based on the experimental results, it is concluded that high-fat feeding during lactation reduces the level of sialylated oligosaccharides in breast milk and modulates the early-life gut microbiota in offspring. These data underline the importance of further research on the effects of early postnatal maternal diet on breast milk composition and the health of offspring.
Author contributions
Lucie Marousez, Marko Verce, Amandine Everard, Delphine Eberlé and Jean Lesage: conceptualization; formal analysis; investigation; project administration; supervision; writing – original draft; writing – review and editing. Laure Dubernat and Thameur Rakza: formal analysis; investigations. Patrice D. Cani, Frédéric Gottrand and Delphine Ley: writing – original draft; writing – review.Data availability
The sequencing data have been submitted to the European Nucleotide Archive (ENA/EBI) and are available under accession number PRJEB83305.Conflicts of interest
PDC is a co-founder of Enterosys SAS (France) and of the Akkermansia company SA. PDC and AE are co-inventors on patents dealing with gut microbes and health.Acknowledgements
This study was supported by grants of the FHU 1000 days for health, the CHRU Lille Hospital and by the French national research program ANR-18-CE21-0005. P. D. C. is an honorary research director at FRS-FNRS (Fonds de la Recherche Scientifique) and recipient of an FRFS-WELBIO grant, WELBIO-CR-2022A-02, and EOS, program no. 40007505. A. E. is a research associate at FNRS (Fonds de la Recherche Scientifique) and recipient of grants from the FNRS under grant no.: J.0075.22 and T.0115.24 and from the FRFS (Fonds de la Recherche Fondamentale Stratégique), with the support of the Walloon region, under grants no.: WELBIO-CR-2019S-03E and WELBIO ADV X.1517.24.References
- C. G.-M. Alcazar, V. M. Paes, Y. Shao, C. Oesser, A. Miltz, T. D. Lawley, P. Brocklehurst, A. Rodger and N. Field, Lancet Microbe, 2022, 3, e867–e880 CrossRef CAS PubMed.
- K. Donald and B. B. Finlay, Nat. Rev. Immunol., 2023, 23, 735–748 CrossRef CAS PubMed.
- C. Milani, S. Duranti, F. Bottacini, E. Casey, F. Turroni, J. Mahony, C. Belzer, S. Delgado Palacio, S. Arboleya Montes, L. Mancabelli, G. A. Lugli, J. M. Rodriguez, L. Bode, W. de Vos, M. Gueimonde, A. Margolles, D. van Sinderen and M. Ventura, Microbiol. Mol. Biol. Rev., 2017, 81, e00036-17 CrossRef PubMed.
- M. Selma-Royo, L. Dubois, S. Manara, F. Armanini, R. Cabrera-Rubio, M. Valles-Colomer, S. González, A. Parra-Llorca, R. Escuriet, L. Bode, C. Martínez-Costa, N. Segata and M. C. Collado, Cell Host Microbe, 2024, 32, 996–1010 CrossRef CAS PubMed.
- H. Szyller, K. Antosz, J. Batko, A. Mytych, M. Dziedziak, M. Wrześniewska, J. Braksator and T. Pytrus, Nutrients, 2024, 16, 1487 CrossRef CAS PubMed.
- C. Lordan, A. K. Roche, D. Delsing, A. Nauta, A. Groeneveld, J. MacSharry, P. D. Cotter and D. van Sinderen, Microbiol. Mol. Biol. Rev., 2024, 88, e0009423 CrossRef PubMed.
- L. Dubernat, L. Marousez, J.-L. Desseyn, V. Gouyer, E. Hermann, F. Gottrand, D. Ley and J. Lesage, Med. Sci., 2023, 39, 869–875 Search PubMed.
- B. Soyyılmaz, M. H. Mikš, C. H. Röhrig, M. Matwiejuk, A. Meszaros-Matwiejuk and L. K. Vigsnæs, Nutrients, 2021, 13, 2737 CrossRef PubMed.
- S. Wu, N. Tao, J. B. German, R. Grimm and C. B. Lebrilla, J. Proteome Res., 2010, 9, 4138–4151 CrossRef CAS PubMed.
- E. Luna, S. G. Parkar, N. Kirmiz, S. Hartel, E. Hearn, M. Hossine, A. Kurdian, C. Mendoza, K. Orr, L. Padilla, K. Ramirez, P. Salcedo, E. Serrano, B. Choudhury, M. Paulchakrabarti, C. T. Parker, S. Huynh, K. Cooper and G. E. Flores, Appl. Environ. Microbiol., 2022, 88, e0148721 CrossRef PubMed.
- A. A. Arzamasov, A. Nakajima, M. Sakanaka, M. N. Ojima, T. Katayama, D. A. Rodionov and A. L. Osterman, mSystems, 2022, 7, e0034322 CrossRef PubMed.
- K. Salli, J. Hirvonen, J. Siitonen, I. Ahonen, H. Anglenius and J. Maukonen, J. Agric. Food Chem., 2021, 69, 170–182 CrossRef CAS PubMed.
- T. Thongaram, J. L. Hoeflinger, J. Chow and M. J. Miller, J. Dairy Sci., 2017, 100, 7825–7833 CrossRef CAS PubMed.
- L. Bode, Nestle Nutr. Inst. Workshop Ser., 2020, 94, 115–123 Search PubMed.
- D. Matharu, A. J. Ponsero, M. Lengyel, A. Meszaros-Matwiejuk, K.-L. Kolho, W. M. de Vos, D. Molnar-Gabor and A. Salonen, EBioMedicine, 2024, 104, 105182 CrossRef CAS PubMed.
- M. B. Azad, B. Robertson, F. Atakora, A. B. Becker, P. Subbarao, T. J. Moraes, P. J. Mandhane, S. E. Turvey, D. L. Lefebvre, M. R. Sears and L. Bode, J. Nutr., 2018, 148, 1733–1742 CrossRef PubMed.
- L. Butruille, L. Marousez, C. Pourpe, F. Oger, S. Lecoutre, D. Catheline, S. Görs, C. C. Metges, C. Guinez, C. Laborie, P. Deruelle, J. Eeckhoute, C. Breton, P. Legrand, J. Lesage and D. Eberlé, Int. J. Obes., 2019, 43, 2381–2393 CrossRef PubMed.
- S. Austin and T. Bénet, Anal. Chim. Acta, 2018, 1010, 86–96 CrossRef CAS PubMed.
- A. Everard, V. Lazarevic, N. Gaïa, M. Johansson, M. Ståhlman, F. Backhed, N. M. Delzenne, J. Schrenzel, P. François and P. D. Cani, ISME J., 2014, 8, 2116–2130 CrossRef CAS PubMed.
- E. Bolyen, J. R. Rideout, M. R. Dillon, N. A. Bokulich, C. C. Abnet, G. A. Al-Ghalith, H. Alexander, E. J. Alm, M. Arumugam, F. Asnicar, Y. Bai, J. E. Bisanz, K. Bittinger, A. Brejnrod, C. J. Brislawn, C. T. Brown, B. J. Callahan, A. M. Caraballo-Rodríguez, J. Chase, E. K. Cope, R. Da Silva, C. Diener, P. C. Dorrestein, G. M. Douglas, D. M. Durall, C. Duvallet, C. F. Edwardson, M. Ernst, M. Estaki, J. Fouquier, J. M. Gauglitz, S. M. Gibbons, D. L. Gibson, A. Gonzalez, K. Gorlick, J. Guo, B. Hillmann, S. Holmes, H. Holste, C. Huttenhower, G. A. Huttley, S. Janssen, A. K. Jarmusch, L. Jiang, B. D. Kaehler, K. B. Kang, C. R. Keefe, P. Keim, S. T. Kelley, D. Knights, I. Koester, T. Kosciolek, J. Kreps, M. G. I. Langille, J. Lee, R. Ley, Y.-X. Liu, E. Loftfield, C. Lozupone, M. Maher, C. Marotz, B. D. Martin, D. McDonald, L. J. McIver, A. V. Melnik, J. L. Metcalf, S. C. Morgan, J. T. Morton, A. T. Naimey, J. A. Navas-Molina, L. F. Nothias, S. B. Orchanian, T. Pearson, S. L. Peoples, D. Petras, M. L. Preuss, E. Pruesse, L. B. Rasmussen, A. Rivers, M. S. Robeson, P. Rosenthal, N. Segata, M. Shaffer, A. Shiffer, R. Sinha, S. J. Song, J. R. Spear, A. D. Swafford, L. R. Thompson, P. J. Torres, P. Trinh, A. Tripathi, P. J. Turnbaugh, S. Ul-Hasan, J. J. J. van der Hooft, F. Vargas, Y. Vázquez-Baeza, E. Vogtmann, M. von Hippel, W. Walters, Y. Wan, M. Wang, J. Warren, K. C. Weber, C. H. D. Williamson, A. D. Willis, Z. Z. Xu, J. R. Zaneveld, Y. Zhang, Q. Zhu, R. Knight and J. G. Caporaso, Nat. Biotechnol., 2019, 37, 852–857 CrossRef CAS PubMed.
- M. Martin, EMBnet J., 2011, 17, 10 CrossRef.
- B. J. Callahan, P. J. McMurdie, M. J. Rosen, A. W. Han, A. J. A. Johnson and S. P. Holmes, Nat. Methods, 2016, 13, 581–583 CrossRef CAS PubMed.
- C. Quast, E. Pruesse, P. Yilmaz, J. Gerken, T. Schweer, P. Yarza, J. Peplies and F. O. Glöckner, Nucleic Acids Res., 2013, 41, D590–D596 CrossRef CAS PubMed.
- M. S. Robeson, D. R. O'Rourke, B. D. Kaehler, M. Ziemski, M. R. Dillon, J. T. Foster and N. A. Bokulich, PLoS Comput. Biol., 2021, 17, e1009581 CrossRef PubMed.
- H. Wickham, M. Averick, J. Bryan, W. Chang, L. D. McGowan, R. François, G. Grolemund, A. Hayes, L. Henry, J. Hester, M. Kuhn, T. L. Pedersen, E. Miller, S. M. Bache, K. Müller, J. Ooms, D. Robinson, D. P. Seidel, V. Spinu, K. Takahashi, D. Vaughan, C. Wilke, K. Woo and H. Yutani, J. Open Source Softw., 2019, 4, 1686 CrossRef.
- P. Paone, D. Latousakis, R. Terrasi, D. Vertommen, C. Jian, V. Borlandelli, F. Suriano, M. E. V. Johansson, A. Puel, C. Bouzin, N. M. Delzenne, A. Salonen, N. Juge, B. I. Florea, G. G. Muccioli, H. Overkleeft, M. Van Hul and P. D. Cani, Gut, 2024, 73, 1632–1649 CrossRef CAS PubMed.
- L. Marousez, L. C. Tran, E. Micours, M. Antoine, F. Gottrand, J. Lesage and D. Ley, Nutrients, 2023, 15, 2771 CrossRef CAS PubMed.
- C. Lozupone and R. Knight, Appl. Environ. Microbiol., 2005, 71, 8228–8235 CrossRef CAS PubMed.
- H. Lin and S. D. Peddada, Nat. Commun., 2020, 11, 3514 CrossRef CAS PubMed.
- K. M. Tonon, M. B. de Morais, A. C. F. V. Abrão, A. Miranda and T. B. Morais, Nutrients, 2019, 11, 1358 CrossRef CAS PubMed.
- M. C. Rudolph, B. E. Young, D. J. Lemas, C. E. Palmer, T. L. Hernandez, L. A. Barbour, J. E. Friedman, N. F. Krebs and P. S. MacLean, Int. J. Obes., 2017, 41, 510–517 CrossRef CAS PubMed.
- B. S. Muhlhausler and G. P. Ailhaud, Curr. Opin. Endocrinol., Diabetes Obes., 2013, 20, 56–61 CrossRef CAS PubMed.
- J. Li, M. Jiang, J. Zhou, J. Ding, Z. Guo, M. Li, F. Ding, W. Chai, J. Yan and X. Liang, Carbohydr. Polym., 2021, 259, 117734 CrossRef CAS PubMed.
- A. Fuhrer, N. Sprenger, E. Kurakevich, L. Borsig, C. Chassard and T. Hennet, J. Exp. Med., 2010, 207, 2843–2854 CrossRef CAS PubMed.
- P. A. Prieto, P. Mukerji, B. Kelder, R. Erney, D. Gonzalez, J. S. Yun, D. F. Smith, K. W. Moremen, C. Nardelli and M. Pierce, J. Biol. Chem., 1995, 270, 29515–29519 CrossRef CAS PubMed.
- F. Liu, A. J. C. Tol, F. Kuipers, M. H. Oosterveer, E. M. van der Beek and S. S. van Leeuwen, Heliyon, 2024, 10, e24539 CrossRef CAS PubMed.
- Y. Wang, B. Rui, X. Ze, Y. Liu, D. Yu, Y. Liu, Z. Li, Y. Xi, X. Ning, Z. Lei, J. Yuan, L. Li, X. Zhang, W. Li, Y. Deng, J. Yan and M. Li, Gut Microbes, 2024, 16, 2334967 CrossRef PubMed.
- M. D. Seferovic, M. Mohammad, R. M. Pace, M. Engevik, J. Versalovic, L. Bode, M. Haymond and K. M. Aagaard, Sci. Rep., 2020, 10, 22092 CrossRef CAS PubMed.
- M. Sakamoto and Y. Benno, Int. J. Syst. Evol. Microbiol., 2006, 56, 1599–1605 CrossRef CAS PubMed.
- R. Schade, L. Song, Z. A. Cordner, H. Ding, D. A. Peterson, T. H. Moran, K. L. Tamashiro and C. B. D. L. Serre, Physiol. Behav., 2023, 258, 113987 CrossRef CAS PubMed.
- P. Louis and H. J. Flint, Environ. Microbiol., 2017, 19, 29–41 CrossRef CAS PubMed.
- K. Borewicz, F. Gu, E. Saccenti, C. Hechler, R. Beijers, C. de Weerth, S. S. van Leeuwen, H. A. Schols and H. Smidt, Sci. Rep., 2020, 10, 4270 CrossRef CAS PubMed.
- J. Astono, Y.-P. Huang, U. K. Sundekilde and D. Barile, Front. Nutr., 2024, 11, 1455251 CrossRef PubMed.
- T. M. Samuel, A. Binia, C. A. de Castro, S. K. Thakkar, C. Billeaud, M. Agosti, I. Al-Jashi, M. J. Costeira, G. Marchini, C. Martínez-Costa, J.-C. Picaud, T. Stiris, S.-M. Stoicescu, M. Vanpeé, M. Domellöf, S. Austin and N. Sprenger, Sci. Rep., 2019, 9, 11767 CrossRef PubMed.
- M. W. Larsson, M. V. Lind, R. P. Laursen, C. Yonemitsu, A. Larnkjær, C. Mølgaard, K. F. Michaelsen and L. Bode, Front. Pediatr., 2019, 7, 297 CrossRef PubMed.
- J. L. Saben, C. R. Sims, A. Abraham, L. Bode and A. Andres, Nutrients, 2021, 13, 446 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d5fo00559k |
| ‡ Co-first authors. |
| § Co-senior authors. |
| This journal is © The Royal Society of Chemistry 2025 |