
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicencePolysaccharides from echinoderms: unlocking health benefits and food applications – a review
Abu
Saeid†
 a,
Deepika
Dave
*ab and
Fereidoon
Shahidi
*a
a,
Deepika
Dave
*ab and
Fereidoon
Shahidi
*a
aDepartment of Biochemistry, Memorial University of Newfoundland, St. John's, NL A1C 5S7, Canada. E-mail: deepika.dave@mi.mun.ca; fshahidi@mun.ca
bMarine Bioprocessing Facility, Centre of Aquaculture and Seafood Development, Marine Institute, Memorial University of Newfoundland, St. John's, NL A1C 5R3, Canada
First published on 24th June 2025
Abstract
Echinoderms (phylum Echinodermata), including sea cucumbers, sea urchins, and starfish, are found in the marine environment. They have no freshwater or terrestrial representatives and inhabit the entire depth of the ocean. The phylum contains more than 7000 living species. Their bodies contain nutrients like proteins (peptides, collagen, and protein hydrolysates), lipids (polyunsaturated fatty acids), saponins (frondoside A), carotenoids (canthaxanthin and astaxanthin), phenolics (flavonoids and phenolic acids), vitamins, and minerals. Besides, these are the leading sources of unique polysaccharides, such as fucosylated chondroitin sulfate, sulfated fucans, and glycosaminoglycans, which possess a wide range of bioactivities. This review intends to explore the health-promoting properties of these polysaccharides, highlighting their anti-inflammatory, anticoagulant, antioxidant, antitumor, anticancer, and other effects along with their mechanisms of action. Their heterogeneous structural composition and remarkable biological activity make them promising candidates for many applications in the functional foods and nutraceuticals area. Furthermore, this review discusses the major challenges and future prospects of polysaccharides from marine echinoderms, focusing on their extraction, purification, characterization, and structural diversity. In addition, the potential of echinoderm polysaccharides as novel nutrients that can contribute to human health is described and it also highlights the growing desire for natural food products in health promotion and disease risk reduction.
 Abu Saeid | Mr Abu Saeid is a PhD student in Food Science at the Department of Biochemistry, Memorial University of Newfoundland, Canada, and a Lecturer (Senior Scale) at Daffodil International University, Bangladesh. He holds a B.Sc. in Food and Process Engineering and an M.S. in Food Processing and Preservation from Hajee Mohammad Danesh Science and Technology University. He has authored over 20 journal articles and 5 book chapters. His research focuses on natural food colorants, marine-derived polysaccharides and protein-peptides, functional foods, seafood processing, and nano-based delivery systems, with additional interests in probiotics, biosensors, bioactive peptides, and active packaging technologies. |
 Deepika Dave | Dr Deepika Dave is a Research Scientist at the Centre for Aquaculture and Seafood Development, Marine Institute, and a cross-appointed Professor in the Department of Biochemistry at the Memorial University of Newfoundland. She holds a PhD in Environmental Engineering from IIT Bombay. Her research focuses on sustainable marine by-product utilization through biorefinery approaches, converting seafood waste into high-value products such as omega-3 fatty acids, chitin, biodiesel, and astaxanthin. Dr Dave has authored multiple peer-reviewed publications on enzymatic hydrolysis, fish oil production, and marine bioactives, contributing to advancing sustainable aquaculture and seafood processing. |
 Fereidoon Shahidi | Dr Fereidoon Shahidi is a University Research Professor and Distinguished Scholar in the Department of Biochemistry at the Memorial University of Newfoundland. He has authored over 1000 research papers and book chapters and edited 80 books, and holds 12 patents. His research focuses on food chemistry, lipid science, antioxidants, phenolics, nutraceuticals, and functional foods. He is the recipient of numerous awards from the Institute of Food Technologists (IFT), American Chemical Society (ACS), American Oil Chemists’ Society (AOCS), Canadian Institute of Food Science and Technology (CIFST), International Union of Food Science and Technology (IUFoST), and International Society for Nutraceuticals and Functional Foods (ISNFF), including lifetime achievement awards from IFT, IUFoST and ISNFF. He serves as the editor-in-chief of the Journal of Food Bioactives and Food Production, Processing and Nutrition; and the founding editor of the Journal of Functional Foods. He has trained many graduate students, postdoctoral fellows, and visiting professors/scholars from over a dozen countries. |
1. Introduction
Echinoderms are marine invertebrates of the phylum Echinodermata, characterized by radial symmetry, a distinctive water vascular (ambulacral) system, and a calcium-rich exoskeleton. They are classified into five major classes (Fig. 1): Asteroidea (starfish), Ophiuroidea (brittle stars), Echinoidea (sea urchins), Holothuroidea (sea cucumbers), and Crinoidea (sea lilies). Echinoderms are exclusively marine, with no freshwater or terrestrial species, and inhabit ocean depths at all levels. The phylum includes over 7000 living species.1,2 Among these, only certain species—particularly sea cucumbers such as Cucumaria frondosa, Apostichopus japonicus, and Stichopus japonicus—are widely consumed for their nutritional and medicinal value. The edible portion is primarily the body wall, while other parts such as viscera/internal organs are generally considered inedible. In contrast, species such as starfish, brittle stars, and sea urchins are largely considered inedible for most of their body weight due to sharp or toxic spines; however, they are valuable sources of bioactive compounds. Nonetheless, certain parts of sea urchins, mainly the gonads, are consumed worldwide. A variety of biomolecules is found in echinoderms, with a diverse range of biological functions. Sea cucumbers contain proteins (peptides, collagen, protein hydrolysates), lipids (polyunsaturated fatty acids), polysaccharides (fucosylated chondroitin sulfate, sulfated fucan), saponins (frondoside A), carotenoids (canthaxanthin and astaxanthin), phenolics (flavonoids and phenolic acids), and minerals.3–10 These components demonstrate diverse bioactivities, including anti-cancer, anti-inflammatory, antioxidant, antithrombotic and anticoagulant, antimicrobial, anti-hypertension, and anti-hyperglycemic activities.11 The structural diversity of polysaccharides in relation to biological function and their qualities in relation to the development of food products such as polysaccharide-based hypoglycemic functional food have drawn much attention in recent years.12,13 | ||
| Fig. 1 Classification of echinoderms (Adapted from Gomes et al.; Arnone et al.; Kalinin and Silchenko).1,2,24 | ||
Sulfated polysaccharides (SPs) and sulfated fucans (SF), collectively known as fucoidan, are important echinoderm polysaccharides. The quantity and ratios of the two SPs differ based on species differences, origin, harvesting time, and the extraction methods employed.14–16 The biological functions of sulfated polysaccharides (SPs) are primarily dictated by their structural characteristics including molecular weight, and the location of the sulfate group. The structures of SPs are primarily influenced by species and extraction methods, exhibiting variations in glycosidic linkage type, sulfation location and degree, sulfate group content, monosaccharide ratios, molecular weight, and other factors.17–20
Until now, several methods have been employed to extract polysaccharides from echinoderms, such as acid and alkaline extraction, microwave-assisted extraction, hydrothermal processing (HP) techniques, high-pressure homogenization (HPH), compressional puffing (CP), ultrasound, β-elimination and oxidative decay techniques, as well as enzyme-mediated depolymerization.8,12,21,22 Compared with plant and fungal polysaccharides, those from aquatic sources are less pigmented and contain fewer molecular contaminants which can be eliminated by continuous precipitation and breakdown.23 Separating SPs is achieved via ethanol precipitation, ammonium salt precipitation, salting out, and ultrafiltration.8 Proteins are among the abundant contaminants in crude polysaccharides extracted from marine species. In recent years, protein elimination and subsequent polysaccharide fraction refining have been carried out. To get rid of polysaccharides from aquatic organisms, chemicals and enzymes, freeze-thaw treatment, adsorption, and salting-out methods have been used.23 Obtaining pure polysaccharides from echinoderms poses various problems, as each process used has its own limitations that need to be considered. As far as we are aware, there is no comprehensive information available on polysaccharides of echinoderms as a whole. Therefore, echinoderm polysaccharides are the focus of this review, along with their structural diversity, biological activity, extraction, purification methods, challenges and proposed potential food applications. The relevant data have been collected from various databases, including Scopus, PubMed, and ScienceDirect, by conducting keyword searches and frequently searching Google Scholar for relevant scholarly literature within 2014 to 2025, but some data were from 2002 to 2013 period.
2. Polysaccharides in marine echinoderms: types and sources
Marine echinoderms (Fig. 2) are among the most leading sources of polysaccharides. Echinoderms have two major classes of fucose-rich sulfated polysaccharides. One type is fucosylated chondroitin sulfate (FCS) found in the body walls of Holothuroidea, such as sea cucumbers, whereas the other is sulfated fucan found in both sea cucumbers and sea urchins.25 Besides these two polysaccharides, sulfated galactans have also been identified in one species of sea urchin. Sea cucumbers, composed of GlcA, GalNAc, Fuc, and sulfate, have high concentrations of FCS. Different sea cucumber FCS's monosaccharide compositions demonstrate approximately equimolar ratios of D-glucuronic acid and D-galactosamine, with a molecular proportion nearing 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1.11 For example, the FCS of Cucumaria frondosa contains D-GalNAc, D-GlcA, L-Fuc, and sulfate in molar ratios of approximately 1:1:1:4.5.26 The FCSs derived from various sea cucumber species may comprise four distinct forms of GalNAc unit—non-sulfated or sulfated at O-4, O-6, or both O-4 and O-6—along with GlcA, which can undergo fucosylation at O-3, sulfation at O-3, or dual sulfation at O-2 and O-3.27,28
:1.11 For example, the FCS of Cucumaria frondosa contains D-GalNAc, D-GlcA, L-Fuc, and sulfate in molar ratios of approximately 1:1:1:4.5.26 The FCSs derived from various sea cucumber species may comprise four distinct forms of GalNAc unit—non-sulfated or sulfated at O-4, O-6, or both O-4 and O-6—along with GlcA, which can undergo fucosylation at O-3, sulfation at O-3, or dual sulfation at O-2 and O-3.27,28
 | ||
| Fig. 2 Marine echinoderms. | ||
Sulfated fucan and sulfated galactan are two high molecular weight polysaccharides commonly extracted from marine invertebrates and other marine sources. They are composed of repeating O-sulfated α-L-fucopyranose (Fucp) or α-L-, α-D-, β-D-galactopyranose (Galp) units with well-defined sulfated patterns, especially those isolated from sea urchins.29,30
Sulfated fucan (SF) from sea cucumbers share many similarities with those found in sea urchins but differ from SF sourced from other marine organisms, such as Phaeophyceae. The most basic structures are polymers composed of 3- or 4-linked mono-sulfated α-L-fucose residues, exemplified by the highly regular polysaccharides [-3)-α-L-Fuc2S-(1-]n identified in Stichopus horrens and Stichopus herrmanni.8,31,32 Sulfated galactans have been identified in two sea urchin species. In Echinometra lucunter, the sulfated galactan consists of α-L-galactopyranosyl units, resembling the polysaccharides in sea squirts. This polysaccharide exhibits greater homogeneity, forming a linear chain of 2-sulfated, 3-linked repetitive units, in contrast to polysaccharides composed of 3-sulfated, 4-linked residues. Another sea urchin species, Glyptocidaris crenularis, contains a galactan composed of 3-linked galactopyranose units in the β-D-enantiomeric form, similar to the galactan found in marine macroalgae.33 In addition, different species of marine starfish contain different polysaccharides. For instance, the polysaccharides glucan and sulfate mannoglucan were isolated from the starfish Asterias rollestoni as fraction 1 and fraction 2, respectively. The main component of fraction 1 was glucan, consisting of 1 → 3 linked β-D-glucopyranose residues. It also contained a minor glucan component with a 1 → 3 linked α-D-glucopyranose residue backbone.34 The mannoglucan sulfate from this species comprises mannose and glucose and contains 13.85% sulfate.35 Different species of brittle star have distinct patterns of sulfation for either chondroitin sulfate or dermatan sulfate. For example, Ramachandra et al. reported chondroitin sulfate/dermatan sulfate from four different species of brittle star, namely Amphiura filiformis, Amphiura chiajei, Ophiothrix fragilis and Ophiocomina nigra. The results showed that Amphiura filiformis had the most tri-O-sulfated disaccharide (2S4S6S), making up about 40% of the sample. The four species also had a considerable amount of 2,4-di-O-sulfated disaccharide units (2S4S), making up about 25–50%.36 Furthermore, a glucan extracted from the brittle star Trichaster palmiferus featured a linear α1,4-linked D-glucopyranose (D-Glcp) backbone, with an α1,4-linked D-Glcp disaccharide side chain attached to the main chain via C-1 to C-6.37
3. Polysaccharide extraction, purification and characterization
Fig. 3 provides an overview of echinoderm polysaccharides’ extraction, purification, and characterization processes. Polysaccharides from echinoderms are extracted through various methods by following several steps. For example, that of sulfated polysaccharides from sea cucumber includes pretreatment, proteolytic digestion, followed by fractionation, and purification. The pretreatment process is the initial step of extraction, where the sample is first eviscerated, and then dried using a dryer. After that, removing fat is carried out, followed by decalcification. Usually, SPs are extracted from fresh sea cucumbers without drying. Different solvent types, including acetone, chloroform, and methanol, are employed during the defatting process of sea cucumber. In comparison with alternative solvents, a 2:1 (v/v) ratio of chloroform to methanol, with or without acetone, showed efficacy in lipid and pigment removal.8 However, the use of non-chlorinated solvents for this purpose is highly recommended. Proteins are one of the major contaminants in sea cucumber polysaccharides coupled with proteoglycans inside their outer layer of cells. For removing these impurities, breakdown of their covalent bond is necessary. Different methods may be employed to break down this bond using enzymes, alkali, hot water or their combinations. Due to increasing solubility, higher dissolution efficacy, and bond (i.e. glycopeptide bonds and other hydrogen bonds) disrupting ability, alkaline hydrolysis with 1–6% NaOH or KOH is recognized as a very common and easy approach. Besides, enzymatic hydrolysis is a popular approach due to its higher extraction efficiency, shorter time of extraction and lower temperature as safe and green methods. A variety of enzymes, including pepsin, trypsin, papain, Alcalase, and Flavourzyme, are employed to extract sea cucumber polysaccharides. For instance, papain at 0.5 to 12.5% was used to isolate GAGs from protein by breaking down their bonds with high efficiency.11 Furthermore, sulfated fucan was extracted using endo-1,3-fucanase to explore its structure, yielding favorable results regarding specificity and enhanced activity.38 Moreover, enzyme-assisted hydrolysis is usually employed to reduce extraction time, increase production yield and then apply alkaline extraction or vice versa. Some emerging technologies like ultrasound, mechanochemical, pulse electric field (PEF), microwave-assisted, enzyme-assisted, high-pressure processing (HPP), and irradiation, among others, have been used for the extraction of SP.39
 | ||
| Fig. 3 Extraction and purification of marine echinoderm polysaccharides. | ||
Various precipitation methods have been utilized for the recovery or separation of glycosaminoglycans, including ethanol, potassium or sodium acetate, quaternary ammonium salts, and the adsorption using an anion exchanger to isolate sulfated polysaccharides from predominantly proteins and other low-molecular-weight contaminants. Ethanol precipitation, typically 40 to 60%, is commonly chosen because of its availability, cost-effectiveness, and lower toxicity. Both chemical (i.e. Sevag and trichloroacetic acid) and enzymatic techniques are often employed for the purification of crude polysaccharides. For non-covalently bonded protein removal, chemical methods, and for covalently bonded protein removal enzymatic methods, have been employed.40 Unlike protein, pigments are among other impurities in crude polysaccharides that need to be removed using organic solvents such as acetone, ethanol and diethyl ether; activated carbon, anion exchange macroporous resin, and hydrogen peroxide may also be employed. However, for removal of inorganic minerals ethanol and water may be used.26,41 For fractionation of the resultant glycosaminoglycans (GAGs), various chromatographic methods, including anion exchange chromatography (employing DEAE Sephacel, DEAE Sephadex, DEAE Sepharose Fast Flow, DEAE-cellulose, Q Sepharose Fast Flow, or strong ion FPA98 resin) and gel-filtration chromatography (utilizing Sephadex, Sephacryl, or Sepharose columns), are often employed. In this, solvents such as ethanol and acetone, membrane filtration, and quaternary ammonium salts have been used.8,40
The next step after purification is characterisation, which is usually done by employing both qualitative and quantitative methods. SPs have been characterized in terms of their monosaccharide composition, structure, sulfation pattern, molecular weight, and methylation. For instance, HPLC may be used to analyze the monosaccharide composition of SPs from sea cucumbers, and gel permeation chromatography (HPGPC) confirmed their purity.42 Nuclear magnetic resonance (NMR) and mass spectrometry (MS) have been employed for structural analysis. It was also confirmed that the entire oligosaccharides had glycosidic bonds as α,β-linkages by observing the correlation signals from their 1D/2D NMR spectra (1H, 13C, COSY, TOCSY, HSQC, ROESY, HMBC). Furthermore, GC-MS, ESI-MS, and FT-IR had been used for the analysis of methylation.42–44
Enzymatic degradation, NMR spectroscopy, methylation analysis, mild acid hydrolysis, sub-critical water processing, oxidative decomposition, and the β-elimination process have frequently been employed to explore the structure of SPs. Enzymatic breakdown yields low molecular mass SPs, leading to a reduced degree of oligosaccharide formation. Under mild reaction conditions, degradation may trigger the breakdown of glycosidic linkages; however, the rate is affected by various parameters including the amount of substrate, temperature, pH, and enzyme concentration.45,46 Mass spectrometry frequently employs mild acid hydrolysis as a preparatory technique for the analysis of oligosaccharides. In the case of FCS, this method selectively removes sulfate groups along with fucose. However, the resulting FCS structure remains susceptible to enzymatic degradation by chondroitinase. The sulfated fucose formed simultaneously and can be identified by NMR spectroscopy. This hydrolysis is effective for qualitatively identifying echinoderms in FCS analysis; however, the extensive removal of sulfate groups from the fragments hinders accurate quantitative assessment.45,47 Methylation profiling is frequently used to accurately identify the positions of glycosidic links and sulfate ester substitutions in polysaccharides. However, the degradation of molecular structures and unexpected changes at the reducing and non-reducing ends of the resulting fragments limit the effectiveness of this method. The trisaccharide fragments α-L-Fucp-(1 → 3)-β-D-GlcA-(1 → 3)-β-D-GalNAc, frequently identified in echinoderms, especially various sea cucumber species, have been obtained using methylation analysis and desulfation. To achieve this desulfation, the sample is subjected to acid hydrolysis, followed by enzyme degradation.11,45 Additionally, oxidative degradation and β-elimination represent green and promising chemical approaches to depolymerise sulfated polysaccharides.8 The FCS core's glycosidic linkage is selectively broken in alkaline conditions by the β-elimination process, resulting in well-defined oligosaccharide structures that boost bioactivity.8,11 Furthermore, the oxidative breakdown of FCS facilitates the synthesis of oligosaccharides through the utilization of H2O2 or Cu+/Cu2+, which produce oxygen free radicals that target glycosidic bonds, especially those involving GalNAc residues. The produced free radical targets the non-sulfated GlcA residues while leaving other glycosidic linkages and sulfate groups intact, leading to the degradation of FCS. Although the oligosaccharide produced by these techniques retains other structural features of the main FCS, their mode of action remains unclear.11,28,46 For instance, Cucumaria japonica FCS has been depolymerized using H2O2 with the inclusion of Cu(OAc)2, leading to a significant reduction in molecular weight while preserving branch and sulfate groups.28 The most popular physicochemical technique for detecting FCS sulfate groups is NMR spectroscopy. Partially depolymerized FCS is valuable for NMR analysis because the original FCS exhibits overcrowded spectra with difficult-to-interpret signals. In contrast to NMR spectroscopy, mass spectrometry (MS), particularly triple MS, is currently employed specifically for structural characterization of SPs; nevertheless, the integration of both techniques may yield reliable results.11 For example, Gao et al. extracted FCS from Holothuria fuscopunctata and determined the distinct structures of several oligosaccharides, including di-, tri-, and tetrasaccharides, produced via mild acid hydrolysis. These structures were further characterized by NMR and MS.44
4. Structural diversity of marine echinoderms polysaccharide
Sulfated polysaccharides, including fucosylated chondroitin sulfate (FCS) and sulfated fucan (SF), are commonly found in echinoderms and show diverse structural variations (Fig. 4). SF from echinoderms consist of repeating tetrasaccharide units, characterized by α(1 → 3) linkages and a consistent sulfation pattern at 2 and 4 positions. Substitutions of sulfo groups are at positions 2-O-, 4-O-, and 2,4-O-. Echinoderms, especially sea urchin polysaccharides such as sulfated fucan and sulfated galactan, consist of α-L-fucopyranoses (Fucp) and either α-L-galactopyranose (Galp), or β-D-Galp oligosaccharides repeating units. However, species-specific variations are observed in certain structural features, including glycosidic linkages [α(1 → 3), α(1 → 4), β(1 → 3)], sulfation patterns at the 2- and/or 4-O positions, and the number of constituent residue units, which can range from tetrasaccharides to disaccharides and monosaccharides, all maintaining a linear configuration. The sulfation patterns and the positions of glycosidic bonds show variability among other species of echinoderms. For instance, the regular repeating sequences of residues in sulfated fucan from sea urchin Lytechinus variegatus are [3-α-L-Fuc2S-1 → 3-α-L-Fuc4S-1 → 3-α-L-Fuc2S4S-1 → 3-α-L-Fuc2S-1]n. However, those in the sulfated fucan from Arbacia lixula sea urchin are [4-α-L-Fuc2S-1 → 4-α-L-Fuc2S-1 → 4-α-L-Fuc-1 → 4-α-L-Fuc-1]n. Additionally, the egg jelly of sea urchins contains galactan sulfate with its oligosaccharides composed of mono- and disaccharide units, for example Echinometra lucunter [→3-α-L-Gal2S-1→]n and Glyptocidaris crenularis [→3-β-D-Gal2S-1 → 3-β-D-Gal-1→]n, respectively.7,48 | ||
| Fig. 4 Linear (a) and branched (b) sulfated fucans from different echinoderms. | ||
Numerous investigations have revealed that SF from the sea cucumber contain linear polysaccharides composed of tetrasaccharide residues of L-fucose linked by (1 → 3) glycosidic linkages and sulfated at the 2 and/or 2,4 positions. For instance, the main structure of sulfated fucan from Holothuria grisea, Holothuria lentiginosa, Holothuria poli, and Holothuria tubulosa was [→3-α-L-Fuc2S-1 → 3-α-L-Fuc2S4S-1 → 3-α-L-Fuc-1 → 3-α-L-Fuc2S-1→]n.49–52 Furthermore, sulfated fucan from Holothuria hilla consisting of highly regular tetrasaccharide repeating units [→3-α-L-Fuc2S-1 → 3-α-L-Fuc2S-1 → 3-α-L-Fuc-1 → 3-α-L-Fuc2S4S-1→]n was obtained, with identical structures to Isostichopus badionotus. This suggests that SF from diverse genera may have comparable core structures.53,54
Holothuria albiventer possesses a unique structural composition of sulfated fucan comprising regular α(1 → 3)-linked uncommon repeating hexasaccharide units.55 Sulfated fucans from the Stichopus genus consist of monosaccharide repeating units. For example, the structure of sulfated fucan [→3-α-L-Fuc2S-1→]n, obtained from Stichopus herrmanni, Stichopus chloronotus, and Stichopus horrens, was established.8,31,32,51
Unlike the (1 → 3) glycosidic linkage, sulfated fucan from different echinoderm species may have other types of glycosidic bond. As an example, Dwivedi et al. identified sea cucumber Thyonella gemmata sequences for sulfated fucan residues as [→3)-α-L-Fuc2S4S-1 → 2-α-L-Fuc4S-1 → 3-α-L-Fuc2S-1 → 3-α-L-Fuc2S-1→] with regular (1 → 2) and (1 → 3)-linked tetrasaccharide repeating units that were sulfated at 2 and 4 positions.56 Additionally, the α-1,4-linked fucans were also identified in Thelenota ananas, Holothuria fuscopunctata, and Pattalus mollis.32,57 Thinh et al. reported that Stichopus variegatus contained sulfated fucan with alternating α(1 → 3) and α(1 → 2) glycosidic bonds.58 Furthermore, a sulfated fucan featuring repeating α(1 → 3) and α(1 → 4) links was derived from Holothuria fuscopunctata.59 A few research studies have shown that the sulfate groups in sulfated fucans could be replaced in locations other than O-2 and (or) O-2,4. For instance, a sulfated fucan from Bohadschia argus is composed of O-4 sulfated fucose residues.28 A sulfated fucan with the structure [→4-α-L-Fuc3S-1→]n was extracted from Holothuria fuscopunctata. Interestingly, the sulfation pattern was located at the O-3 position, which is relatively rare in sulfated fucan, as most glycosidic linkages occur at the O-4 position.59
The linear structure of sulfated fucan is predominantly observed in diverse echinoderms, such as sea cucumbers, while several species exhibit fucose branching. For instance, Apostichopus japonicus was identified to have a branched sulfated fucan consisting of a new pentasaccharide repetitive unit represented as [→3-α-L-Fuc2S-1 → 3, (α-L-Fuc-1 → 4-α-L-Fuc-1 → 4-α-L-Fuc2S-1 → 3-α-L-Fuc2S-1→]n.
Besides, sulfated fucan containing a fucose branch has been obtained from Holothuria edulis.18 Additionally, this sulfated fucan consists of typical α(1 → 4)-linked tetrasaccharide repetitive units and 4-O-sulfated fucose side chains derived from Holothuria coluber.42 Sometimes sulfated fucan comprises hexasaccharide repetitive units with an α(1 → 3) linkage and is sulfated at different positions. For instance, sulfated fucan extracted and purified from Holothuria albiventer consists of hexasaccharide repeating units with α(1 → 3) linkages and sulfation at various positions, including O-3, O-3,4, O-2,3, or O-2,4.55
Fucosylated chondroitin sulfate (FCS) is typically composed of equimolar amounts of α-L-Fuc, β-D-GlcA, and GalNAc units, with its core consisting of α(1 → 3) or alternate α(1 → 3) and α(1 → 4) connected fucose residues.25 Echinoderms, especially sea cucumbers, contain FSC abundantly consisting of glucuronic acid (GlcA), N-acetyl-D-galactosamine (GalNAc), fucose (Fuc), and sulfate (Table 1). The monosaccharide composition of different sea cucumber FCS shows about equivalent amounts of D-GlcA and D-GalNAc, with a molecular ratio of around 1:1.11 FCS has been extracted from over 30 species of sea cucumber. The standard chondroitin core of this polysaccharide is →4)-β-D-GlcpA-(1 → 3)-β-D-GalpNAc-(1→, which is sulfated at the O-4 and/or O-6 position of GalNAc, and contains sulfated α-L-fucosyl residues as branches linked to the O-3 of GlcA in the backbone.60 For instance, Thyonella gemmata FCS backbone consists of [→3)-β-GalNAc-(1 → 4)-β-GlcA-(1→] comprising 70% 4-sulfated and 30% 4,6-disulfated GalNAc units, with one-third of the GlcA residue infused at the C3 position by branching α-fucose (Fuc) molecules, which are either 65% of 4-sulfated or 35% 2,4-disulfated.56 It is essential to mention that the actual diversity of natural FCS structures is considerably broader (Fig. 5). For example, GlcA units, both fucosylated and sulfated at O-3, were identified in multiple fucosylated chondroitin sulfates.26 Fucosyl branches linked to both GlcA and GalNAc units have been proposed for FCS derived from Apostichopus japonicus,61 and confirmed for FCS from Actinopyga mauritiana.26 A study indicated that di-fucosyl residues may replace the O-3 of GlcA in conjunction with mono-fucosyl units,26 but this structural characteristic was not demonstrated. GalNAc sulfation differed in degree among species of sea cucumbers.18 The FCS extracted from Ludwigothurea grisea comprised 53% of 6-sulfated GalNAc units, while the polysaccharide derived from Stichopus japonicus had all 4,6-disulfated GalNAc residues.26
 | ||
| Fig. 5 Structural diversity of fucosylated chondroitin sulfate from echinoderms. | ||
| Echinoderm species | Mole ratio (%) | Molar mass (kDa) | Ref. | ||||
|---|---|---|---|---|---|---|---|
| GlcA | GalNAc | Fuc | Gal | Sulfate | |||
| GlcA, glucuronic acid; GalNAc, N-acetyl-D-galactosamine; Fuc, fucose, and Gal, galactose. | |||||||
| Cucumaria japonica | 1 | 0.73 | 1.38 | — | 4.5 | 32 | 28 |
| Bohadschia argus | — | 0.78 | 1 | 0.09 | 4.04 | 32 | 28 |
| Holothuria (Theelothuria) spinifera | — | 0.74 | 1 | 0.12 | 3.44 | 30 | 28 |
| Lysastrosoma anthosticta | — | 22.7 | — | 0.5 | 24.8 | 12.5 | 71 |
| Stichopus japonicus | 1 | 1.07 | 1.21 | — | 3.71 | 60.99 | 27 |
| Holothuria hilla | 13.3 | — | 10.8 | 1.6 | 21.2 | — | 64 |
| Hemioedema spectabilis | 1.1 | 1 | 1.15 | — | 3.9 | 44.1 | 64 |
| Holothuria fuscopunctata | 1 | 1.18 | 1 | — | 33.7 | 42.6 | 59 |
| Stichopus japonicus | 0.9 | 1 | 3.1 | 0.2 | 3.5 | 224.2 | 67 |
| Stichopus chloronotus | 0.9 | 1 | 1.08 | — | 3.18 | 111 | 41 |
| Acaudina molpadioidea | 0.82 | 1 | 0.88 | — | 3.04 | 93.3 | 41 |
| Apostichopus japonicus | 0.98 | 1 | 1.15 | — | 3.65 | 98.1 | 41 |
| Holothuria tubulosa | 12 | 14 | 17 | — | 57 | 54.02 | 31 |
| Acaudina molpadioidea | 0.82 | 1 | 0.88 | — | 3.04 | 93.3 | 41 |
| Cucumaria djakonovi | 16 | 16 | 11 | — | 57 | 17.3 | 31 |
| Cucumaria frondosa | 12 | 12 | 14 | — | 62 | 24.02 | 60 |
| Thelenota ananas | 1 | 1.02 | 0.98 | — | 3.81 | 60.9 | 72 |
| Isostichopus badionotus | 1 | 0.7 | 0.9 | — | 3.1 | 109 | 73 |
| Pearsonothuria graeffei | 1 | 0.8 | 1.5 | — | 2.6 | 73 | 73 |
| Actinopyga mauritiana | 17 | 18 | 13 | — | 52 | 26.43 | 26 |
| Strongylocentrotus nudus | 0.21 | 0.12 | 0.63 | 0.29 | — | — | 74 |
| Glyptocidaris crenularis | 0.62 | 0.58 | 0.43 | 1.21 | — | — | 74 |
| Anthocidaris crassispina | 0.14 | 0.06 | 0.08 | 0.25 | — | — | 74 |
| Asterias rollestoni | 1 | — | — | — | 13.85 | 151.1 | 34 |
FCS's molecular weight varies, ranging from 30 to 100 kDa, based on the methods employed for extraction. For instance, when extracting Apostichopus japonicus FCS using enzymes, the molecular weight was 27 kDa (ref. 26) but other researchers found higher values when using combinations of alkali and enzymes.41,43
Additionally, several studies have demonstrated that FCS from identical species, including Apostichopus japonicus and Stichopus chloronotus, exhibit variations in molecular weight.62 Chondroitin sulfate (CS) from marine invertebrates is categorized as CS-A (GlcA-GalNAc4S), CS-C (GlcA-GalNAc6S), CS-D (GlcA2S-GalNAc6S), CS-E (GlcA-GalNAc4S6S), and CS-O (GlcA-GalNAc) units according to the sulfation pattern.63 Numerous sea cucumber species, including Cucumaria japonica, Cucumaria frondosa, Eupentacta fraudatrix, and Hemioedema spectabilis, predominantly demonstrate sulfation at the O-3 position of GlcA residues. The prevalent disaccharide residue combinations are GlcA3S-GalNAc4S6S, GlcA3S-GalNAc4S, GlcA3S-GalNAc, and GlcA2S3S-GalNAc6S.26,60,64 The sulfation patterns of fucose branches vary depending on the species and/or preparation method. The patterns of fucose side chains (Table 2) include non-sulfated, 3-sulfated, 4-sulfated, 2,4-disulfated, or 3,4-disulfated forms.65 For instance, Mansour et al. identified a Fuc2,4S and Fuc3,4S branching FCS from Holothuria poli.66 Three different sulfated fucose residue types—Fuc2,4S, Fuc3,4S, and Fuc4S—were found in FCS from Stichopus japonicus at a ratio of 2:1.5:1.67 Generally, Fuc monosaccharides linked to the O-3 of GlcA formed the branches in FCS, but various analytical methods also reported the emergence of new branches and connection types. For instance, FCSs from Holothuria mexicana contained fucosyl residues linked to O-6 and O-4 of GalNAc and multi-fucosyl branches linked to O-3 of GlcA.68 Even in the same species, such as Apostichopus japonicus, FCS structures were different.61,69
| Echinoderms species | Fuc0S | Fuc3S | Fuc4S | Fuc2S4S | Fuc3S4S | Ref. |
|---|---|---|---|---|---|---|
| The branching Fuc unit can be sulfated at the 2, and/or 3 and/or 4-positions according to the species. | ||||||
| Bohadschia argus | 0 | 0 | 0 | 5 | 95 | 28 |
| Holothuria mexicana | — | — | 3.03 | 1 | — | 75 |
| Apostichopus japonicus | 0 | 0 | 17 | 54 | 29 | 43 |
| Holothuria mexicana | 6.24 | 5.58 | 37.16 | — | 51 | 68 |
| Apostichopus japonicus | — | — | 0.26 | 1 | 0.71 | 41 |
| Acaudina molpadioidea | — | — | 3.99 | 1 | — | 41 |
| Stichopus chloronotus | — | — | 0.24 | 1 | — | 41 |
| Holothuria tubulosa | — | — | 14.3 | 42.85 | 42.85 | 31 |
| Holothuria stellati | — | — | 20 | 40 | 40 | 31 |
| Holothuria scabra | — | — | 27.1 | 72.9 | — | 42 |
| Cucumaria djakonovi | — | — | ∼25 | ∼50 | ∼25 | 31 |
| Actinopyga mauritiana | — | — | — | 20 | 80 | 31 |
| Holothuria coluber | 0 | 21 | 21 | 32 | 26 | 42 |
| Cucumaria japonica | — | — | — | 20 | 80 | 31 |
| Cucumaria frondosa | — | — | — | 25 | 62.5 | 60 |
| Holothuria poli | 0 | 0 | 0 | 46 | 54 | 66 |
| Eupentacta fraudatrix | 33.3 | — | — | — | 66.7 | 60 |
| Apostichopus mauritania | — | — | — | 16.7 | 66.7 | 26 |
| Pattalus mollis | 0 | 0 | 26 | 34 | 40 | 17 |
| Holothuria forskai | — | — | 15 | 39 | 46 | 76 |
| Holothuria nobilis | 52.94 | 35.29 | 0 | 11.77 | 0 | 77 |
| Ludwigothurea grisea | 0 | 0 | ∼49 | ∼20 | ∼17 | 25 |
| Acaudina molpadioidea | 40.3 | 0 | 11.94 | 23.88 | 23.88 | 77 |
| Thelenota ananas | 0 | ∼25 | ∼22 | ∼55 | 0 | 78 |
| Stichopus tremulus | — | — | 24.8 | 22.4 | 52.8 | 69 |
| Pearsonothuria graeffei | — | — | 81.6 | 18.4 | — | 69 |
| Isostichopus badionotus | — | — | 4.1 | 95.9 | — | 69 |
In addition to other polysaccharides, echinoderms contain glucans. For example, the brittle star species Trichaster palmiferus possesses a novel glucan with a linear α-(1 → 4)-Glcp backbone and disaccharide side chains of α-D-Glcp-(1 → 4)-α-D-Glcp-(1→), which are linked to every fourth Glcp unit at the C-6 position in the backbone.37 Furthermore, Castro discovered that the egg jelly coat of sea urchins contains sulfated galactan.70
The structural diversity of polysaccharides from marine echinoderm is closely tied to the species and tissue type—particularly distinguishing between edible and inedible components. Thus, sea cucumber species such as Cucumaria frondosa and Apostichopus japonicus, along with other species, primarily yield fucosylated chondroitin sulfate (FCS) and sulfated fucan (SF) from their edible body wall, with well-characterized sulfation patterns and defined molecular structures that contribute to their safety and efficacy as food-derived functional ingredients. In contrast, some echinoderms like sea urchins and brittle stars often contain structurally distinct polysaccharides, such as sulfated galactans or dermatan sulfates, which are typically extracted from shells, gonads or whole-body tissue. These polysaccharides often reveal structural heterogeneity, unique sulfation sites, and larger molecular weight ranges, lending better themselves to pharmaceutical or cosmeceutical applications with limited food use.7,9–11,36,48 Therefore, comprehending the structural differences between polysaccharides from edible and inedible sources is essential for directing their use in targeted nutritional, therapeutic, or industrial applications.
5. Marine echinoderm polysaccharides in health promotion
Echinoderms, a diverse group of marine invertebrates, encompass sea cucumbers, sea urchins, starfish, brittle stars and sea lilies, each playing a vital role in ocean ecosystems. They are the major echinoderms containing biologically active compounds, especially bioactive polysaccharides that have potential health benefits (Fig. 6). Researchers have summarized the therapeutic potential of these compounds, highlighting their antioxidant, anti-inflammatory, anti-thrombotic, anti-tumor, and anti-cancer properties (Table 3). | ||
| Fig. 6 Health benefits of echinoderm polysaccharides. | ||
| Health effects | Marine echinoderms | Tissue type and edibility | Polysaccharides | Mode of action | Ref. |
|---|---|---|---|---|---|
| a Does not clearly mention the body parts. | |||||
| Anti-cancer | Cucumaria frondosa | Body wall (edible) | Fucosylated chondroitin sulfate | Reduced the aggregation and cytotoxicity of Aβ40 | 8 |
| Cucumaria frondosa | Body wall (edible) | Sulfated fucan | Stop human osteosarcoma epithelial cells from sticking to fibronectin and stopping them from migrating; slow down the cytoskeleton's ability to change shape by stopping the phosphorylation of focal adhesion kinase and paxillin | 99 | |
| Hemioedema spectabilis | Body wall (edible) | Fucosylated chondroitin sulfate | Blocked cancer cells from sticking to the surface covered with platelets and suppressed the formation of tubulogenesis | 64 | |
| Stichopus japonicus | Extracta | Sulfated fucan | Sensitize colorectal cancer cells to TRAIL-induced apoptosis by promoting XIAP ubiquitination and inducing endoplasmic reticulum stress | 98 | |
| Isostichopus badionotus | Fragmentsa | Fucosylated chondroitin sulfate | LMW sulfated fragments inhibited A549 lung cancer cell growth and migration more | 87 | |
| Stichopus japonicus | Extracta | Sulfated fucan | By producing NK cells and cytotoxicity | 100 | |
| Cucumaria frondosa | Body wall (edible) | Fucosylated chondroitin sulfate | Inhibited the signaling cascade of the ERK1/2/p38 MAPK/NF-κB axis, which was crucial for the production of MMPs | 65 | |
| Strongylocentrotus nudus | Egg (edible) | Polysaccharide | SEP suppressed lung cancer proliferation by stimulating NK and T cells through TLR2/4, driving the release of potent effectors like perforin and IFN-γ by the activated immune cells | 135 | |
| Asterina pectinifera | Extracta | Starfish polysaccharides | Increase the activity of TIMP-1, which stops MMP-9 from working; lower the activity of cells that metastasize; and stop MMP-9 from working in MCF-7 human breast cancer cells | 107 | |
| Anti-coagulation | Acaudina leucoprocta | Body wall (edible) | Fucosylated chondroitin sulfate | Prolonging APTT, TT and PT and inhibition of AT III-mediated F II and FX | 118 |
| Cucumaria frondosa | Body wall (edible) | Fucosylated chondroitin sulfate | Prevent Xase complex formation by attaching the negatively charged sulfate group of FCS to the positively charged side chain of arginine residues on the specific surface of factor IXa | 114 | |
| Stichopus naso | Body wall (edible) | Sulfated fucan/fucosylated glycosaminoglycan | Prolonging APTT activity and iXase inhibition | 117 | |
| Isostichopus badionotus | Body wall (edible) | Fucosylated chondroitin sulfate | MW reduction greatly lowered APTT, TT, and IF-Xase | 56 | |
| Holothuria fuscopunctata | Body wall (edible) | Fucosylated glycosaminoglycan | By prolonging APTT activities | 136 | |
| Ludwigothurea grisea | Body wall (edible) | Fucosylated chondroitin sulfate | An inhibitory effect on Xase complex formation | 113 | |
| Stichopus monotuberculatus | Body wall (edible) | Fucosylated glycosaminoglycan | Block the intrinsic tenase complex selectively | 115 | |
| Bohadschia argus Holothuria spinifera | Body wall (edible) | Fucosylated chondroitin sulfate | Strengthen thrombin's inhibitory efficacy by establishing a ternary complex with antithrombin III | 28 | |
| Holothuria floridana | Body wall (edible) | Sulfated fucan | By prolonging APTT activities | 121 | |
| Holothuria nobilis | Body wall (edible) | Fucosylated chondroitin sulfate | Extended APTT and prevented intrinsic coagulation Xase | 8 | |
| Pattalus mollis | Body wall (edible) | Sulfated polysaccharides | APTT prolongation and intrinsic factor Xase prevention | 57 and 120 | |
| Holothuria hilla | Body wall (edible) | Fucosylated chondroitin sulfate | The presence of ATIII inhibits thrombin and factor Xa | 64 | |
| Cucumaria syracusana | Body wall (edible) | Fucosylated chondroitin sulfate | Mediation of heparin cofactor II (HCII) | 116 | |
| Holothuria fuscopunctata | Body wall (edible) | Sulfated polysaccharides | Inhibit the forming of Xase | 59 | |
| Paracaudina chilensis | Body wall (edible) | Fucosylated chondroitin sulfate | Inhibited the formation of thrombin and factor Xa in the presence of AT III | 64 | |
| Acaudina leucoprocta | Body wall (edible) | Sulfated fucan | Selective anti-thrombin activity facilitated by heparin cofactor II | 119 | |
| Holothuria poli | Body wall (edible) | Fucoidan | Inhibit formation of thrombin, reduction of platelet aggregation | 19 and 51 | |
| Apostichopus japonicus | Body wall (edible) | Fucosylated glycosaminoglycan | Hindering action on Xase complex formation | 43 | |
| Holothuria scabra | Body wall (edible) | Fucosylated chondroitin sulfate | By prolonging activated partial thromboplastin time | 42 | |
| Holothuria albiventer | Body wall (edible) | Sulfated fucan | Increased extension of APTT and suppression of intrinsic factor Xase production | 55 | |
| Stichopus horrens | Body wall (edible) | Fucosylated glycosaminoglycan | Suppression of the intrinsic coagulation cascade through selective inhibition of human intrinsic factor Xase | 32 | |
| Holothuria mexicana | Body wall (edible) | Fucosylated chondroitin sulfate | Intrinsic anticoagulant activity through AT III-mediated suppression of thrombin and factor Xa production | 68 | |
| Thelenota ananas, Holothuria edulis, Holothuria nobilis, Holothuria fuscopunctata, Stichopus herrmanni | Body wall (edible) | Fucosylated chondroitin sulfate | Thrombin inhibition via heparin cofactor II activation | 86 | |
| Holothuria poli | Body wall (edible) | Fucosylated chondroitin sulfate | Activity is mainly governed by HCII and, to a lower degree, by antithrombin (AT) | 66 | |
| Cucumaria japonica | Body wall (edible) | Fucosylated chondroitin sulfate | Platelet aggregation inhibition by collagen and ristocetin | 137 | |
| Anti-inflammation | Holothuria tubulosa | Extracta | Sulfated fucan | Suppress inflammatory cytokine production and render M2 polarization in co-cultured macrophages through PPARγ activation | 20 |
| Cucumaria frondosa, Holothuria albiventer | Body wall (edible) | Sulfated fucan/fucosylated chondroitin sulfate | Inhibits proinflammatory cytokines, including TNF-α and IL-6 production | 13 | |
| Isostichopus badionotus | Body wall (edible) | Fucosylated chondroitin sulfate | Inhibits TPA-induced overexpression of TNF-α, IL-6, NF-κB, iNOS, IL-10, IL-11, COX-2, and STAT3 in mouse ear tissue at the genetic level | 83 | |
| Acaudina molpadioides | Body wall (edible) | Fucosylated chondroitin sulfate | Reduce the levels of LPS and TLR4 signaling. Enhancing the levels of fecal SCFAs triggers the activation of AMPK, ultimately resulting in the development of inflammation resistance | 130 | |
| Apostichopus japonicus | Body wall (edible) | Sulfated fucan | By suppressing the MAPK/NF-κB and AKT/mTOR signaling pathways and downregulating iNOS expression in the livers of lipopolysaccharide-challenged mice | 85 | |
| Apostichopus japonicus, Stichopus chloronotus | Body wall (edible) | Fucosylated chondroitin sulfate | Reduces neutrophil infiltration and mitigates paw edema | 41 | |
| Paracentrotus lividus | Eggs (edible) | Sulfated polysaccharide | Lower the paw swelling | 88 | |
| Anthocidaris crassispina | Shell (inedible) | Shell polysaccharide | Reduces nitric oxide (NO) production | 74 | |
| Antidiabetic | Stichopus japonicus | Body wall (edible) | Fucosylated chondroitin sulfate | Facilitated dose-dependent absorption of glucose and utilization in insulin-resistant HepG2 cells | 27 |
| Cucumaria frondosa, Thelenota ananas | Body wall (edible) | Fucosylated chondroitin sulfate | In T2DM rats, it enhances the IRS/PI3K/AKT signaling cascade and modulates GSK-3β activity | 13 | |
| Holothuria leucospilota | Body wall (edible) | Polysaccharide | Mitigates the clinical manifestations of T2DM in Goto-Kakizaki rats | 127 | |
| Cucumaria frondosa | Body wall (edible) | Fucosylated chondroitin sulfate | - Ameliorated endoplasmic reticulum stress and hepatic inflammation in mice with high-fat sugar diet (HFSD) induced insulin resistance | 81 and 128 | |
| - Suppressed cell death in pancreatic islets by blocking the mitochondrial pathway | |||||
| Anti-viral | Isostichopus badionotus | Body wall (edible) | Fucosylated chondroitin sulfate and Sulfated fucan | Antiviral effects that inhibit adenovirus and human cytomegalovirus by preventing viral attachment and cellular infiltration | 138 |
| Patallus mollis | Extracta | Sulfated glycans | Anti-viral activity against human rotavirus | 139 | |
| Thelenota ananas | Body wall (edible) Extracta | Sulfated polysaccharides/fucosylated chondroitin sulfate | - Inhibit SARS-CoV-2 host cell entrance via binding to the S glycoprotein | 133 and 134 | |
| - Inhibiting the reproduction of multiple HIV-1 variant and preventing viral adhesion and cellular infiltration by targeting the gp120 protein | |||||
| Anti-oxidant | Stichopus chloronotus | Body wall (edible) | Fucoidan | This new fucoidan markedly decreased the degradation of lipids | 51 |
| Stichopus chloronotus, Apostichopus japonicus, Acaudina molpadioidea | Body wall (edible) | Fucosylated chondroitin sulfate | The presence of sulfate residues may contribute to the scavenging of free radicals | 41 | |
| Acaudina molpadioidea and Holothuria nobilis | Body wall (edible) | Fucosylated chondroitin sulfate | Exhibit dose-dependent moderate antioxidant action over DPPH, hydroxyl, and superoxide radicals | 140 | |
a. Anti-inflammatory properties
Inflammation is a contributing factor to damage generated by metabolic syndrome. Anti-inflammatory agents can reduce inflammation. Marine echinoderms possess biologically active components, such as polysaccharides, that have been shown to exhibit anti-inflammatory properties, highlighting their potential for therapeutic applications. Numerous investigations have demonstrated that polysaccharides extracted from sea cucumbers exhibit anti-inflammatory effects. A diverse array of sea cucumbers, including Cucumaria frondosa, Isostichopus badionotus, Thelenota ananas, Stichopus variegatus, Stichopus (Apostichopus) japonicus, Holothuria nobilis, Stichopus choloronotus, and Ypsilothuria bitentaculata, are well studied. Sulfated fucan (SF), fucoidan, and fucosylated chondroitin sulfate (FCS) are the most prevalent polysaccharides commonly extracted, all of which show anti-inflammatory properties in different animal and cellular models.79 FCS from sea cucumbers has been found to exert anti-inflammatory effects in different insulin-resistant models. For instance, FCS from Apostichopus japonicus decreased the levels of IL-1β, TNF-α, and IL-6 in the livers of high-fat diet (HFD) fed mice.80 Additionally, FCS from Cucumaria frondosa reduced serum TNF-α levels in an HFD-induced mouse model of insulin resistance.13 Furthermore, fucosylated chondroitin sulfate had diversified targeted functions in the reduction of inflammation. For example, FCS from Cucumaria frondosa suppressed kappa kinase beta (IKKβ) and c-Jun NH2-terminal kinase (JNK), effectively limiting NF-κB nuclear translocation in the liver of HFD-induced insulin-resistant mice, thereby mitigating pro-inflammatory signaling.81 Mou et al. demonstrated that FCS significantly reduced carrageenan-induced swelling in a dose-dependent manner within an inflammatory model.41 FCS isolated from edible sea cucumbers, such as Apostichopus japonicus, Stichopus chloronotus, Cucumaria djakonovi, and Acaudina molpadioidea, effectively alleviated carrageenan-induced paw edema in a mouse model.31,41 SF- or FCS-enriched polysaccharide fraction from the low-edible sea cucumber species was found to lower the pro-inflammatory cytokine levels (including TNF-α and IL-6) in HFD or streptozotocin (STZ) treated rats, leading to a decrease in inflammation in these rats.13 Sulfated polysaccharide (heteroglycan) fractions extracted from the body parts of Cucumaria frondosa impaired cell survival, as reflected by the increased level of TNF-α, IL-6, and IL-10 in THP-1 macrophages.82 Purified FCS from Isostichopus badionotus inhibited NF-κB gene expression, suppressing downstream targets such as COX-2, iNOS, and TNF-α. Consequently, this reduced inflammation and cell damage induced by 12-O-tetradecanoylphorbol-13-acetate in a mouse ear inflammation model.83 Long-term administration of fucoidan from Isostichopus badionotus lowered hepatic expression and serum levels of inflammatory cytokines and biomarkers, including TNF-α, IL-1β, IL-6, IL-10, macrophage inflammatory protein 1 (MIP-1), and C-reactive protein obese mice induced by diet. Because fucoidan turned off the JNK and IκB/NF-κB pathways in hepatocytes, it was able to reduce inflammation.84 Furthermore, fucoidan extracted from Apostichopus japonicus attenuated the expression of pro-inflammatory factors (TNF-α, IL-1β, and IL-6) by preventing the MAPK/NF-κB signaling cascade in a mouse model of liver injury induced by lipopolysaccharide (LPS).85 Additionally, fucoidans from Thelenota ananas stopped stomach ulcers caused by alcohol by lowering the levels of proinflammatory cytokines and related transcription factors like TNFα, IL-6, and NF-κB.86 Furthermore, fucoidan oligosaccharides from Pearsonothuria graeffei and Isostichopus badionotus helped reduce low-grade inflammation caused by an HFD in mice by reducing the concentrations of TNFα and LPS in their blood.87 Zhu et al. reported that sulfated fucan isolated from Holothuria tubulosa has the capability to activate PPARγ, decrease the synthesis of inflammatory cytokines, and enhance M2 polarization in co-cultured macrophages.20 Moreover, high molecular weight is sometimes responsible for reducing inflammation. For example, sea urchin (Paracentrotus lividus) eggs possess a sulfated polysaccharide of higher molecular weight that mitigates carrageenan-induced edema in rat paws. This polysaccharide works by stopping the production of or blocking the effects of different chemicals like serotonin, histamine, prostanoids, and leukotrienes.88 A study reports shell and gonad polysaccharides worked directly or indirectly against inflammation by inhibiting the cell growth. For example, Jiao et al. indicated that the polysaccharides found in the shell and gonads of the purple-colored sea urchin showed cytotoxicity towards RAW 264.7 cells, which are implicated in inflammation.74b. Antioxidant properties
The body, under stress, generates reactive oxygen species (ROS) which are associated with various diseases, including inflammatory disorders, cancer, aging, cardiovascular diseases (CVD), neurological dysfunction, and cataracts.5,89 Antioxidants can inhibit free radical generation, diminish oxidative stress within cells, and provide a protective effect against illnesses generated by ROS. Thus, dietary antioxidants may play a role in preventing disorders caused by oxidative stress, and hence natural food products containing antioxidants have garnered interest, particularly in view of the detrimental impact of synthetic antioxidants.90 As a result, echinoderms that possess bioactive substances, including polysaccharides with antioxidant effects, are important. The antioxidant activity of marine echinoderm polysaccharides has been assessed using in vitro assays, including hydroxyl, ABTS, DPPH, ROS scavenging, superoxide radical scavenging, and reducing power.34 Sulfated fucan (SF) found in sea cucumbers is employed as a dietary supplement because of its well-known antioxidant properties. Sulfated fucan is a prospective antioxidant capable of inhibiting free radicals that induce cellular damage in the body. Li et al. isolated SF from Stichopus chloronotus, which contained L-fucose and sulfate esters. This polysaccharide significantly inhibited lipid peroxidation in RAW 264.7 cells and lowered NO and TNF production.51 Increasing the sulfate level and decreasing the molecular weight of polysaccharides like SF improves their antioxidant activity, thereby preventing superoxide radical formation.11 SF derived from Thelenota ananas at a concentration of IC50 17.46 μg mL−1 effectively reduced superoxide radicals, with the sulfation pattern significantly influencing this activity.91 SF from Holothuria fuscopunctata also demonstrated antioxidant activity against superoxide radicals; however, it is ineffective against ABTS, DPPH or hydroxyl radicals. This dissimilarity is probably attributed to the structural properties of SF and the distinct mechanisms of action employed by the different in vitro assays.44 Fucosylated chondroitin sulfate (FCS) exhibits potent antioxidant activity. A study showed that the polysaccharide GAG derived from Holothuria mexicana effectively scavenges DPPH, superoxide, and hydroxyl radicals in a dose-dependent manner.41 The degree and specific sites of sulfation influence FCS's antioxidant properties.92 This is because the polysaccharides contain many hydroxyl and reducing groups. For example, FCS derived from three edible sea cucumbers—Acaudina molpadioidea, Apostichopus japonicus, and Stichopus chloronotus—exhibited superior antioxidant potential, driven by distinct sulfation patterns that enhance bioactivity. At a concentration of 4 mg mL−1, these species demonstrated remarkable nitrate radical scavenging activity, ranging from 26% to 39%.41,93 Investigations have shown that sulfated polysaccharides from Holothuria fuscogilva possess strong antioxidant properties. The results indicated that the EC50 values for hydroxyl and superoxide radical scavenging activity of Holothuria fuscogilva were 3.74 mg mL−1 and 0.037 mg mL−1, respectively.72In vivo and in vitro research has shown that FCS from sea cucumber reduces oxidative stress. For example, FCS derived from Stichopus chloronotus protected the intestinal barrier and improved antioxidant function in H2O2-injured Caco-2 cells by upregulating tight junction proteins and triggering the Keap1-Nrf2-ARE antioxidant pathway. Furthermore, giving FCS to mice that had been treated with cyclophosphamide (Cy) could stop them from losing weight and having a smaller spleen. It could also increase the levels of ZO-1, Claudin-1, Nrf2, SOD, and NQO-1 in Cy-damaged colon tissue, protecting against oxidative stress and damage to the intestinal barrier in vivo.94 An unknown polysaccharide from sea cucumbers (Stichopus japonicus Selenka) gonad showed higher antioxidant activity when it was extracted using an ultrasound-assisted enzymatic extraction. Degradation of the gonad polysaccharide chains led to the exposure of more sulfate groups through the breaking of glycosidic bonds within the polysaccharide. Sea cucumber gonad polysaccharides from 0.25 to 4.00 mg mL−1 demonstrated higher DPPH, OH, and ferric reducing/antioxidant efficacy.95 Research suggests that starfish contain polysaccharides with significant antioxidant properties. For example, a glucan and sulfate mannoglucan were isolated from the starfish species Asterias rollestoni, referred to as fractions SF1 and SF2, respectively. SF2 had the highest superoxide scavenging activity effect because of the highest sulfate contents, and the IC50 concentration was 4 mg mL−1 as compared with SF1. Conversely, the SF2 fraction demonstrated 100% hydroxyl radical-scavenging activity at a concentration of 10 mg mL−1.34c. Anticancer activity
Continuous and excessive cell division causes cancer. These diseases typically undergo evolution through somatic cell mutations, with a small number of mutations being inherited, arising randomly, or resulting from exposure to mutagens. Cancer pathogenesis is primarily attributed to mutations in oncogenes, which dominate in promoting growth over tumor suppressor genes and the dysfunction of these suppressor genes.89,96,97 Anticancer properties from natural sources, especially marine sources, have been reported for echinoderm polysaccharides, particularly fucosylated chondroitin sulfate (FCS) and sulfated fucan (SF). SF demonstrates potential in anticancer therapy. For example, sulfated fucan from Stichopus japonicus may make colorectal cancer (CRC) cells more sensitive to TRAIL therapy. This occurs through inhibiting the X-linked inhibitor of apoptosis protein (XIAP) ubiquitination and suppressing endoplasmic reticulum (ER) stress.98 Additionally, studies have demonstrated that sulfated fucan from Cucumaria frondosa inhibits the adhesion of human osteosarcoma epithelial cells to fibronectin and suppresses their motility.99 Furthermore, sulfated fucan derived from Stichopus japonicus was isolated and separated into different fractions, namely F1, F2, F3, and F4, in order to examine their ability to activate natural killer (NK) cells and induce cytotoxicity. The F1 treatment showed the most potent NK cell cytotoxicity (45% at 250 g mL−1) against HeLa cells, while the F3 and F4 treatments showed strong NK cell cytotoxicity (31–34% at 250 g mL−1) against HepG2 and HT-29 cells.100 Studies have demonstrated that low molecular weight FSC derived from Cucumaria frondosa suppresses the development and metastasis of Lewis lung carcinoma (LLC) in a dose-dependent manner. The process by which FCS caused cell cycle arrest in LLC cells involves raising p53/p21 expression and triggering apoptosis by activating caspase-3 activity.65 FCS from Apostichopus japonicus was administered to mice in a dose-dependent manner, resulting in increased cytotoxicity of NK cells to YAC-1 cells and CTL cells to B16 cells.101 Holothurian glycosaminoglycan (hGAG) is extractable from the body wall of the sea cucumber. The treatment of lung cancer cells, specifically lung adenocarcinoma A549 cells, involves the combined administration of hGAG and cisplatin (DDP). DDP exerts an inhibitory effect on A549 lung cells through the apoptosis pathway.102 Liu et al. demonstrated that polysaccharides from Strongylocentrotus nudus eggs exhibit a notable preventive impact on mouse tumor cells and also discovered that the polysaccharide D-glucan was the cause of tumor cell death.103 One of the polysaccharides found in Strongylocentrotus nudus eggs, SEP, demonstrated anti-hepatocellular carcinoma activity. These polysaccharides show great potential as therapeutic candidates for liver cancer treatment. In the same way, in vitro conditions revealed that a polysaccharide from sea urchin gonads had anti-tumor activity.104 In addition, a soluble polysaccharide isolated from the gut of sea urchins greatly hindered the development of human gastric tumor cells (SGC-7901) and hepatoma cells (Bel-7402), with the suppression of growth increasing in a concentration-dependent manner.105 With an unidentified composition, crude polysaccharides from echinoderms have demonstrated notable anti-cancer activity. The polysaccharide derived from starfish shows potential as a chemopreventive agent for human breast cancer. As an example, polysaccharides from the starfish Asterina pectinifera, despite lacking detailed structural characterization, have been shown to inhibit the proliferation of human breast cancer cell lines, including estrogen receptor-positive (MCF-7) and estrogen receptor-negative (MDA-MB-231) cells.106 By decreasing the level of COX-2 and MMP-9, which are controlled by an MAPK signaling pathway, this might stop cancer cells from spreading and getting worse in MCF-7 cells.107 In the same way, starfish polysaccharide stops the development and metastasis of MDA-MB-231 breast cancer cells by lowering the expression of COX-2 and aromatase and stopping the cells from moving.108 In addition, the polysaccharide from Asterina pectinifera species demonstrated protective effects against colon cancer by inducing detoxification enzymes and inhibiting the activities of ODC and COX-2.109 Furthermore, the anticancer activity of brittle stars was demonstrated for the crude polysaccharide separated from the Persian Gulf brittle star Ophiocoma erinaceus, which inhibited the development of HeLa cells in a dose- and time-dependent manner. This isolated polysaccharide induced intrinsic apoptosis by increasing caspase-3, caspase-9, and Bax levels while decreasing Bcl-2 levels in HeLa cells.110 Moreover, brittle star polysaccharides exhibited anti-angiogenic activity. For instance, a polysaccharide isolated from brittle stars demonstrated a strong anti-angiogenic effect by reducing VEGF levels. It also showed anti-metastatic effects associated with inhibiting MMP-9 and paxillin expression.111d. Anticoagulant and anti-thrombotic properties
Thromboembolic disorders have a significant impact on mortality and disability rates worldwide.8 Uncontrolled blood clotting and heightened micro-thrombus production are primary factors leading to organ malfunction and mortality in affected individual.112 Anticoagulating agents are crucial for individuals with a high susceptibility to blood clots since they effectively reduce the likelihood of experiencing severe health complications such as strokes and heart attacks.89 Hence, foods that include anticoagulant attributes can effectively mitigate the danger of thromboembolic ailments. Marine echinoderms have a myriad of secondary metabolites, like fucosylated chondroitin sulfate and fucoidan, that have good anticoagulant properties. Apart from fucosylated chondroitin sulfate (FCS), the level of anticoagulant activity of this polysaccharide usually varies based on the fucose branches in its structure. For example, FCS were isolated from four species of sea cucumber. Two of these sea cucumbers contained sulfated fucose (SF) branches in their structures and exhibited the strongest anticoagulant activity with values of 135 (APTT assay), and 132 IU mg−1 (TT assay) for Stichopus tremulus, as well as 183 (APTT assay), and 157 IU mg−1 (TT assay) for Isostichopus badionotus.69 FCS with low molecular weight (5.3 kDa) from Ludwigothurea grisea exhibits increased anti-coagulation activity, primarily by inhibiting the formation of the Xase complex, and shows promising anticoagulant properties.113 In addition, low molecular weight (10 kDa) FCS from the sea cucumber Cucumaria frondosa was shown to possess increased intrinsic anti-coagulant activity. The study demonstrated that FCS binds its negatively charged sulfate group to the positively charged arginine residues on a specific surface of factor IXa, thereby preventing the formation of the Xase complex.114 The anticoagulant characteristics of fucosylated glycosaminoglycan, extracted from Apostichopus japonicus, have been reported.43 These findings show that Apostichopus japonicus exhibits intrinsic factor Xase inhibition in the absence of any adverse side effects, including platelet aggregation and FXII activation. The anticoagulation activities of fucosylated glycosaminoglycan has also been demonstrated. The authors reported that fucosylated glycosaminoglycan selectively inhibits intrinsic tenase complexes while not impacting the activity of anti-factor IIa, Xa, and VIIa under the examined conditions.115 Chahed et al. demonstrated that FCS derived from Cucumaria syracusana exhibited significant anticoagulation activity, primarily attributed to heparin cofactor II (HCII) and, to a limited extent, antithrombin (AT), with IC50 values of 0.05 μg mL−1 for HCII and 0.09 μg mL−1 for AT.116 Moreover, Cui et al. demonstrated the anticoagulant properties of sulfated fucan and fucosylated glycosaminoglycan isolated from Stichopus naso. These findings suggest that oligosaccharides with a lower degree of polymerization, particularly octasaccharides and 2,3-disulfated fucose (Fuc2S4S), lead to a more significant prolongation of APTT and enhanced iXase inhibition.117 The anticoagulation and antithrombotic action of Acaudina leucoprocta FCS has been reported, demonstrating a strong antithrombin III (AT III) binding affinity. This interaction effectively reduces the activities of factor IIa (F IIa) and factor Xa (F Xa), resulting in anticoagulant and antithrombotic effects.118Several reports have suggested that sulfated fucan from sea cucumber has anticoagulant activities. For example, sulfated fucan from Acaudina leucoprocta exhibits selective anti-thrombin activity through the modulation of heparin cofactor II, employing a mode of action distinct from heparin.119 Additionally, Zheng et al. (2019) demonstrated the anticoagulation actions of two sulfated polysaccharides isolated from Pattalus mollis, both of which significantly reduced activated partial thromboplastin time (APTT) and the intrinsic factor IXa–VIIIa complex (Xase), leading to prolonged effects.57 Zheng et al. and Ma et al. found five Pattalus mollis sulfated fucans with anticoagulant activities. Their research findings showed that Pattalus mollis sulfated fucans exhibit substantial and dose-dependent anticoagulant properties.57,120 Like Pattalus mollis, sulfated polysaccharides extracted from Holothuria fuscopunctata also demonstrated inhibitory activity against the intrinsic factor Xase complex.59 A strong APTT-prolonging activity was also observed in the SF obtained from Holothuria floridana.121 Additionally, Mansour et al. (2019) documented the anticoagulant effects of fucoidan (1376 kDa), which was extracted from Holothuria poli. A calibrated automated thrombography approach was used to measure thrombin production. The findings indicated that the isolated polysaccharide significantly inhibited thrombin production in a dose-dependent manner, showing a significant effect at 5 μg mL−1 under the conditions tested.19 Moreover, Li et al. (2020) demonstrated the anticoagulant effects of a fucoidan (103.1 kDa) derived from Holothuria poli utilizing a Balb/c mouse model. They suggested that fucoidan diminishes platelet aggregation induced by cyclophosphamide, a chemotherapeutic agent with extensive anticancer efficacy.51 Most of the time coagulation is linked to platelet aggregation, as with heparin, but few studies have suggested anticoagulant action without platelet aggregation. For example, Wu et al. identified a novel polysaccharide in sea cucumbers Holothuria edulis and Ludwigothurea grisea. The structures comprised a primary backbone made up of common α(1 → 3)- and α(1 → 2)-linked tetrasaccharide repeating units as a side chain of an unsulfated fucose residue. The results demonstrated that FS showed potent anticoagulant activity. This activity was attributed to its antithrombin activity through heparin cofactor II, without inducing platelet aggregation. It is crucial to underscore that the anticoagulant mechanisms of sulfated fucan differ from those of heparin-like agents.18 In addition to the studies mentioned, the 3-linked 2-sulfated galactan from the sea urchin Echinometra lucunter and the 3-linked 2-sulfated fucan from Strongylocentrotus franciscanus have also been found to inhibit thrombin activity.122 According to Sibiya et al. a mix of sulfated glycans from various sea urchins included 3-linked 4-sulfated fucan from Lytechinus variegatus, 3-linked 2-sulfated fucan from S. franciscanus, and a 3-linked 2-sulfated galactan from Echinometra lucunter.123
The structurally related sulfated glycans mentioned above have potential as antithrombotic or anticoagulant drugs and could be administered orally or in gastroresistant tablet form.124
e. Anti-diabetic properties
Diabetes mellitus, especially type-2, is a prevalent and chronic metabolic disorder characterized by elevated high blood glucose levels.125 Insulin resistance or reduced insulin secretion in the pancreas leads to decreased blood insulin activity, which accelerates lipid peroxidation, contributing to diabetes and its associated complications. A prolonged high glucose level in the blood, called hyperglycemia, leads to oxidative stress, inflammation, impaired glucose tolerance and other complications.126 Consequently, it results in clinical symptoms associated with diabetes. Insulin resistance and hyperglycemia are the two key characteristics of type 2 diabetes mellitus (T2DM). Numerous research studies have indicated that polysaccharides derived from sea cucumbers demonstrate anti-diabetic properties. For instance, fucosylated chondroitin sulfate (FCS) from Cucumaria frondosa was administered to mice in a high-fat diet (HFD) dose-dependent manner, improving insulin resistance and anti-hyperglycemic effects.13Holothuria leucospilota (HLP) polysaccharides significantly worsened glucose intolerance and influenced blood lipid and hormone concentrations.127 Gong et al. indicated that chondroitin sulfate derived from Stichopus japonicus exhibits anti-diabetic properties. The study suggested that the polysaccharides enhance glucose consumption and absorption in insulin-resistant HepG2 cells in a dose-dependent manner, potentially mitigating cellular damage.27 Additionally, they promoted glycogen production in glucosamine-induced insulin-resistant HepG2 cells. FCS derived from Cucumaria frondosa enhances insulin sensitivity and insulin resistance, as well as repairs pancreatic islet apoptosis, in vivo.39 Hu et al. suggest that FCS in Acaudina molpadioides enhances glucose metabolism.128Sulfated fucan (SF) has demonstrated significant antihyperglycemic effects. An example is SF from Cucumaria frondosa, which can greatly lower insulin and blood sugar levels in mice that are insulin-resistant by turning on the PI3K/PKB pathway and GLUT4 in fat and muscle.129 SF obtained from Acaudina molpadioides improved the hyperglycemia and insulin resistance in in vivo studies.128,130Acaudina molpadioides, Isostichopus badionotus, Thelenota ananas, Holothuria tubulosa, and Pearsonothuria graeffei are some of the sea cucumber species that produce sulfated fucose. This polysaccharide has strong effects on improving insulin resistance. The findings showed that fucose with a lower molecular weight and O-4-sulfation substitution can effectively reduce insulin resistance.131
f. Miscellaneous health benefits
Research has demonstrated remarkable hypolipidemic activity of sulfated fucan (SF). Li et al. proposed that SF derived from Pearsonothuria graeffei can mitigate gaining weight in high-fat diet (HFD) mice. Additionally, it may ameliorate hyperlipidemia and safeguard the liver against lipid breakdown.68 Moreover, SF from Thelenota ananas demonstrated anti-adipogenic effects by modulating adipocyte development and lipid deposition.6 Furthermore, SF derived from Acaudina molpadioides exhibits a notable inhibitory effect on lipogenesis. For instance, mice administered sulfated fucan (200 μg mL−1) exhibited a 34.07% lessening in fat beneath the skin, around the reproductive organs, and within their cells compared with those fed a HFD.132 The anti-hyperlipidemic properties of fucoidans derived from Pearsonothuria graeffei and Isostichopus badionotus have been previously reported. Comparative analysis indicates that the sulfated fucan from P. graeffei exhibits superior anti-hyperlipidemic activity, as evidenced by the downregulation of CYP7A1 and the upregulation of PPARα. These molecular changes affect fatty acid transport into hepatic cells and control the metabolism of lipid, emphasizing the potential of P. graeffei-derived fucoidan in lipid-lowering applications. On the other hand, Isostichopus badionotus exhibited only minor effects.72 Fucoidan derived from Acaudina molpadioides shows significant anti-adipogenic effects via influencing the Wnt/β-catenin pathway and decreasing the expression of SREBP-1c.132 Furthermore, fucosylated chondroitin sulfate exhibits anti-hyperlipidemic properties. For example, FSC extracted from Stichopus japonicus showed a dose-dependent inhibitory action on pancreatic lipase.67 In addition, FCS from Isostichopus badionotus and Pearsonothuria graeffei demonstrated considerable efficacy in improving lipid profiles, regulating glucose levels in the blood, protecting liver function, and enhancing hepatic glycogen synthesis.73 More recently, the anti-obesity properties of echinoderm polysaccharides have been reported. For instance, administration of Stichopus japonicus to HFD-fed mice led to reduced body weight, enhanced insulin sensitivity, decreased adipose and hepatic enlargement, and lower serum lipid levels and inflammatory cytokine concentrations.16Several studies have shown that sulfated fucans and fucosylated chondroitin sulfate derived from echinoderms enhance gut microbiota regulation and promote microbial community diversification. For example, sulfated fucan derived from Stichopus japonicus promoted the growth of short-chain fatty acid-producing bacteria and sulfide-degrading bacteria while declining the harmful bacteria population.16 Li et al. highlighted the impact of fucoidan derived from Pearsonothuria graeffei on gut microbial dysbiosis and metabolic disorders in HFD-fed mice. The findings demonstrated that the polysaccharide effectively decreased weight gain, relieved hyperlipidemia, and safeguarded the liver against steatosis in HFD mice.68 Thus, gut microbiota can convert Thelenota ananas sulfated fucan into short-chain fatty acids, thus increasing Bacteroidetes and Firmicutes and decreasing Proteobacteria.52 Furthermore, Fuc-Pg increased the prevalence of Bacteroidetes and Actinobacteria while diminishing the quantity of Firmicutes and Proteobacteria. An in vivo study revealed that chondroitin sulfate derived from Isostichopus badionotus influences the gut microbiota composition.87 FCS derived from sea cucumbers mitigated gut microbiota dysbiosis by enriching bacterial diversity and reshaping the microbial community structure. For example, FCS isolated from Stichopus chloronotus increased the proportion of beneficial microbiota while suppressing the proliferation of pathogenic bacteria.94
Research indicates that fucosylated chondroitin sulfate (FCS) exhibits antiviral properties. Specifically, depolymerized FCS from Thelenota ananas (TadFCS, 12.5 kDa with a CS-O/A backbone) effectively blocked human immunodeficiency virus (HIV) entry and replication. In particular, Ta-dFCS has shown the ability to inhibit the multiplication of many HIV-1 strains in vitro, encompassing laboratory strains (HIV-1IIIB and HIV-IRF), clinical strains (HIV-1KM018 and HIV-1 TC-2), as well as T-20-resistant strains.133 Additionally, sulfated polysaccharides (FCS and SF) from the sea cucumber Stichopus japonicus showed promising inhibitory effects against SARS-CoV-2 transmission, with an IC50 value of 9.10 μg mL−1.134 Fucosylated chondroitin sulfate and sulfated fucan from Thyonella gemmata were found to inhibit SARS-CoV-2 pseudovirus-carrying spike proteins from both the Delta (B.1.617.2) and wild-type (Wuhan-Hu-1) strains. Notably, SF exhibited greatly enhanced anti-SARS-CoV-2 activity against both variants.56
6. Application in the food industry
For decades, food manufacturing companies have had an interest in polysaccharides owing to their varied functional properties, serving as stabilizers, thickening agents, emulsifiers, and humectants.141 Currently, the food industry utilizes an abundance of polysaccharides including agar, alginates, gum Arabic, carrageenan, methylcellulose, xanthan, dextran, and pectin. Such compounds are derived from animal, plant, marine, and microbial origins. Marine-derived polysaccharides’ chemical composition and structural characteristics are key to their bioactivity. This marine-derived polysaccharide possesses several attributes that render it a suitable food ingredient. Fucan/fucoidan, carrageenan, hydrocolloids, and glycosaminoglycans are polysaccharides typically utilized across various industries, particularly in the food, beverage, and supplement sectors. Seaweed, crustaceans, and other marine species are the immediate sources of these polysaccharides.142 For example, algae-derived polysaccharides function as gelling agents, emulsifiers, clarifiers, thickeners, stabilizers, and flocculants in ice cream, candies, yoghurt, meat products, and functional beverages. They are also available as alginate, agar, and carrageenan.143 The distinctive structural properties of this polysaccharide, including its gelling and thickening abilities, result from polysaccharide–polysaccharide interactions. Agar's ability to form gels makes it widely applicable in dairy products, baked goods, canned fish and meat, sauces, soups, beverages, and various culinary preparations, including traditional Japanese dishes like “Tokoroten” (gelled agar strands) and “Mitsumame” (a dessert with sweet syrup, fruits, and agar cubes).144 In addition, due to high sugar content, agar is employed in confectionary productions particularly fruit candy, custards, jams, jellies, and puddings. Like agar, alginate is utilized for the manufacturing of jams and puddings due to its firm and rigid gel-forming capabilities. Additionally, it serves as a thickener in ketchup, ice cream, sauces, purees and mayonnaise. In addition, pastries and fruit fillings use alginate's ability to form films to keep the cake from becoming moist.142 Researchers have reported on the water binding and dispersing properties of polysaccharides. These properties alter the water-binding and foaming attributes of food products, thereby modifying their texture.145 Carrageenan, employed in dairy-related products such as processed cheeses, flans, sterilized chocolates, and evaporated milk, had similar properties.142 Chitosan, along with other marine polysaccharides, are very suitable as raw materials for the production of edible and biodegradable films. They can also be used as antibacterial agents, additives, and nutraceuticals. Furthermore, polysaccharides such as carrageenan, chitosan, and chito-oligosaccharides can be added to meat products, sausages, dairy products, and beverages.5,39,146,147 In addition, fucoidan from Acaudina molpadioides, a sea cucumber species, has been incorporated into multilayer emulsions having caseinate-coated lipid droplets, enhancing stability in various food matrices.49 However, echinoderm, especially sea cucumber-related, products are available, but no studies have summarized the prospect of sea cucumber polysaccharides producing functional foods. Unlike polysaccharides from other marine sources, echinoderm polysaccharides demonstrate significant potential for application in the development of functional foods. Fig. 7 demonstrates the proposed approach to developing functional food using echinoderm polysaccharides, which can reduce non-communicable diseases.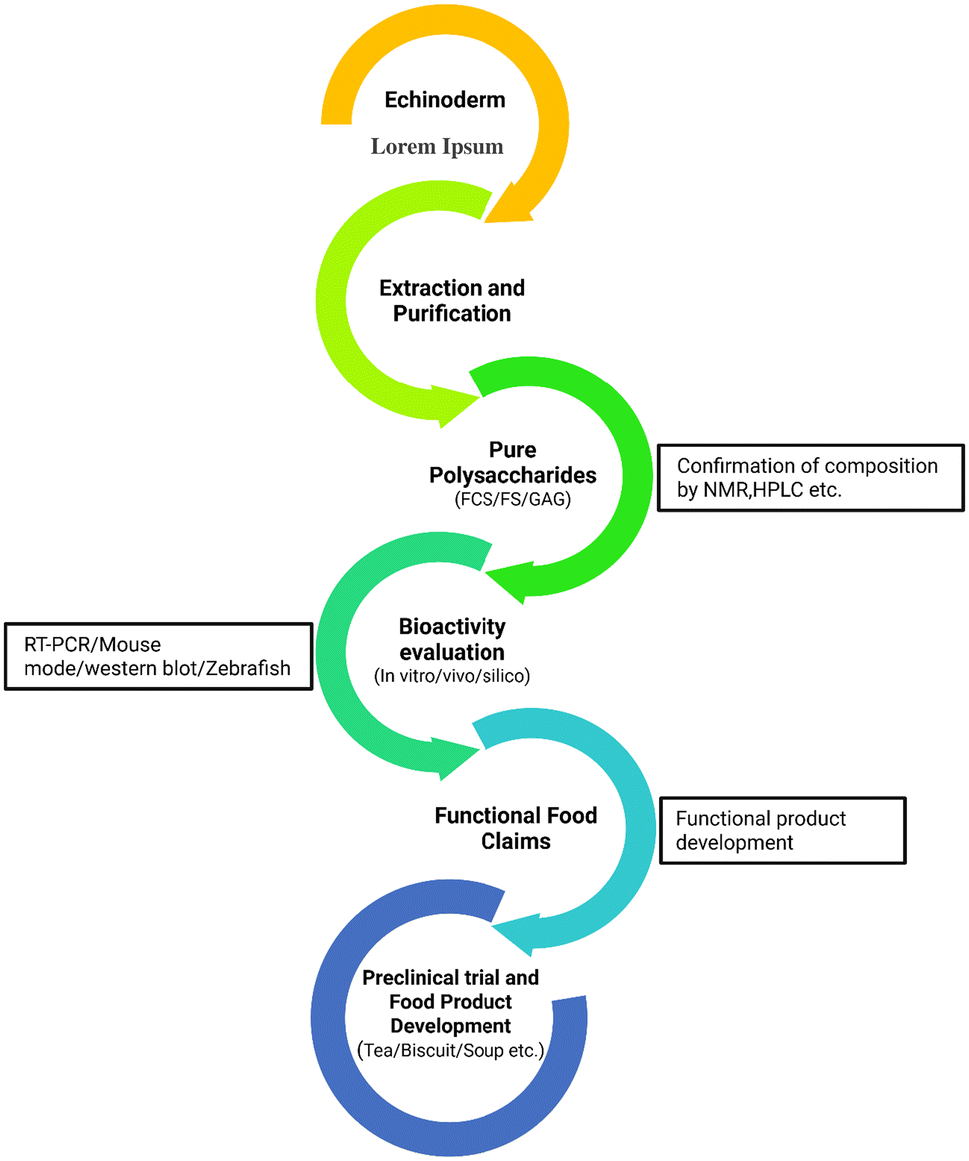 | ||
| Fig. 7 Proposed approach for developing functional food from echinoderms. | ||
7. Barriers and breakthroughs in echinoderm polysaccharides
Polysaccharides derived from marine sources, particularly marine echinoderms, exhibit a distinctive structure that enables various physiological and therapeutic actions. These compounds are gaining popularity worldwide because of their widespread availability, simple extraction process, unique structures, minimal adverse effects, and extensive pharmacological applications.23 However, their journey to market remains incomplete. The structural features of these polysaccharides greatly affect their bioactivities, making it essential to employ appropriate extraction and purification methods to retain their bioactivity.a. Challenges in extraction methods
Currently, numerous extraction techniques are used to isolate crude polysaccharides, but the choice of method greatly impacts the structural integrity of sulfated polysaccharides (SPs). Acids and alkalis are commonly employed to disrupt protein interactions and free SPs.61 However, these chemicals can alter SP structures, with alkalis having a more pronounced effect, including the breakdown and removal of sulfate groups.23,77,148 Similarly, techniques like trichloroacetic aid (TCA) precipitation and calcium chloride salting out can break glycosidic bonds, thus further complicating the extraction process.13 Advanced technologies, although not without limitations, offer alternatives to traditional methods. For instance, microwave-assisted extraction can reduce molecular weight and cleave sulfate groups.149,150 Another promising approach, hydrothermal processing (HP), enhances SP yields while preserving sulfate groups, although it is time-consuming.21,151b. Impurities
Impurities such as pigments and proteins present additional challenges. In aquatic animals, polysaccharides are often bound to proteins as glycoproteins, making their removal difficult. Various approaches, including chemical and enzymatic techniques, have been employed to extract proteins. The Sevag method, while commonly used, is inefficient and requires repeated operations, leading to polysaccharide loss.152 The TCA method is more effective but causes glycosidic bond cleavage and hazardous solvent residues, rendering the resulting products unsuitable for pharmaceutical and food applications.13,153 Resin absorption offers another option for protein removal but results in higher polysaccharide loss.154 Recently, a freeze–thaw method has emerged as a novel technique with higher polysaccharide recovery rates, though it is time- and energy-intensive.c. Structural diversity and analysis
A significant hurdle in utilizing these polysaccharides lies in their structural diversity. Variability in polysaccharide structures, even within the same species of echinoderms like sea cucumbers, complicates functional studies. Structural examination must precede any biological study to ensure reproducibility and clarity. Structural analysis of heterogeneous fragments also requires further refinement. Sensitive and effective methodologies for polysaccharide structural investigation are crucial. While online sequencing methods for proteins and nucleic acids are well-established, contemporary techniques for polysaccharide analysis, such as NMR, are still offline and time-consuming. The abundance of sulfate groups further complicates structural determination.155,156d. Global context and future directions
The growing global demand for marine polysaccharides in pharmaceuticals, functional foods, and cosmetics underscores the importance of addressing these challenges. Developing scalable, eco-friendly extraction methods while maintaining the structural integrity of SPs is vital. Innovations such as enzyme-based extractions, non-invasive purification technologies, and automated structural analysis tools could significantly enhance commercial viability. Collaborative actions between researchers, industry stakeholders, and policymakers will be indispensable for unlocking the full potential of echinoderm polysaccharides across diverse applications.e. Future prospects and research directions
The efficacy of echinoderm polysaccharides, including fucosylated chondroitin sulfate (FCS) and sulfated fucan (SF), as integral elements of functional foods and medicinal agents has been well established. Products derived from echinoderms, such as sea cucumber polysaccharides (a combination of SF and FCS), have already entered the market. In 2023, the global chondroitin sulfate market reached an estimated value of $1.29 billion, with an anticipated compound annual growth rate (CAGR) of 3.6% (2024 to 2030). This growth rate reflects the market's steady growth caused by rising demand for nutraceuticals, osteoarthritis treatments, and pharmaceuticals. North America led the market (∼$484.7M), while the Asia-Pacific region is projected to experience a faster growth trajectory, with a CAGR of 4.2% over the forecast period. Bovine-derived chondroitin sulfate held the largest share (38%), with dietary supplements dominating applications (35%). The market is poised for expansion due to rising consumer awareness and pharmaceutical applications. Chondroitin sulfate is initially extracted from animal sources such as bovine cartilage, shark cartilage, and pig trachea, with bovine-derived chondroitin sulfate being the most widely used in commercial products.157,158 However, chondroitin sulfate from echinoderms remains mainly unexplored in the market and presents potential for future innovation. Developing and commercializing products featuring high-purity, structurally defined FCS and SF—or those explicitly labelled as such—continues to be an unmet objective. Achieving this milestone is critical to maximizing the nutritional and therapeutic potential of these compounds for consumers.To advance the application of FCS and SF in medicine and nutrition, several areas require further exploration and development. Comprehensive toxicological studies, pharmacokinetic evaluations, and the production of innovative drug delivery systems, such as nanoparticles and hydrogels, are essential to expand their medical use, particularly in wound healing, tissue engineering, and regenerative medicine. The proangiogenic and anti-inflammatory properties of FCS and SF highlight their potential in wound healing and tissue engineering applications. These compounds can enhance tissue regeneration and repair by promoting new blood vessel formation and modulating inflammation. For example, research involving zebrafish larvae demonstrated that FCS extracted from green and purple sea cucumbers (Apostichopus japonicus) enhanced caudal fin regeneration. The treatment increased fin regrowth area and improved locomotor behaviours within 48 hours post-amputation. Additionally, there was an upregulation in the expression of genes associated with regeneration, including Wnt10a, msx1b, fgf20a, bmp2a, bmp4, and igf2b. The study also observed elevated mRNA levels of inflammatory factors and genes associated with the TLR/NF-κB signaling pathway.159 This suggests that FCS may facilitate tissue regeneration by modulating inflammatory responses and triggering distinct signaling pathways. A study investigated the effects of fucoidan on wound healing. The findings suggest that fucoidan promotes angiogenesis through the AKT/Nrf2/HIF-1α signaling pathway, expediting wound healing. Moreover, it supports the proliferation of hydrogen peroxide (H2O2)-damaged human umbilical vein endothelial cells (HUVECs) and enhances endothelial cell network development. Mechanistic studies revealed that fucoidan enhanced the protein expression of the AKT/Nrf2/HIF-1α signaling pathway, a key regulator of angiogenesis. This effect was further validated by the inhibitor LY294002, which counteracted fucoidan's ability to promote endothelial tube network formation.160 While precise studies directly linking these polysaccharides to such applications (wound healing and tissue engineering) are limited, echinoderms polysaccharides especially FCS and SF are well-known but their remarkable regenerative abilities are only now the subject of extensive research. Additionally, examining the antiviral properties of FCS and SF against emerging diseases offers promising research opportunities.
Another important avenue is the investigation of the synergistic effects of FCS and SF when combined with chemotherapeutic agents. This could lead to the formulation of targeted anticancer therapies, potentially enhancing treatment efficacy and patient outcomes. Developing scalable production methods for these polysaccharides while maintaining their structural integrity and bioactivity will be pivotal in realizing their full potential as functional and therapeutic agents.
8. Conclusion
Marine echinoderms are a powerhouse of dietary polysaccharides, especially fucosylated chondroitin sulfate and sulfated fucan. The bioactivity of these polysaccharides varies with their structural diversity, particularly the position of sulfate groups and fucose content. Distinguishing between edible and inedible echinoderm sources is critical when evaluating their polysaccharides for nutritional and therapeutic relevance. Edible sea cucumbers provide body wall-derived polysaccharides with promising applications in functional foods and nutraceuticals, supported by their safety and long-standing consumption history. In contrast, polysaccharides from inedible sources—such as sea urchin shells and brittle stars—are structurally distinct and show potential in pharmaceutical and biomedical applications, though they may not be suitable for food use. Recognizing and addressing these differences enables more targeted utilization of echinoderm polysaccharides and better alignment with specific industry needs.This review focuses on the extraction, purification, and structural characterization of echinoderm-derived polysaccharides, emphasizing their bioactivities—particularly anticancer, anti-inflammatory, antidiabetic, anticoagulant, and antioxidant properties. Their incorporation into functional food formulations is also discussed. This contribution desires to support researchers and product developers in the sustainable and purposeful exploitation of this invaluable marine resource by addressing existing challenges and identifying future research directions.
Author contributions
Abu Saeid: conceptualization, methodology, investigation, and writing – original draft; Fereidoon Shahidi and Deepika Dave: supervision, validation, writing – reviewing and editing, and funding acquisition.Conflicts of interest
There are no competing interests to declare.Data availability
No additional data are available.Acknowledgements
This research was funded by the Natural Sciences and Engineering Research Council (NSERC) of Canada.References
- V. I. Kalinin, Echinoderms Metabolites: Structure, Functions, and Biomedical Perspectives, Mar. Drugs, 2021, 19, 1–5 CrossRef PubMed.
- A. R. Gomes, A. C. Freitas, T. A. Rocha-Santos and A. C. Duarte, Bioactive compounds derived from echinoderms, RSC Adv., 2014, 4, 29365–29382 RSC.
- T. R. Senadheera, A. Hossain, D. Dave and F. Shahidi, Functional and physiochemical properties of protein isolates from different body parts of North Atlantic sea cucumber (Cucumaria frondosa), Food Biosci., 2023, 52, 1–10 CrossRef.
- T. R. Senadheera, A. Hossain, D. Dave and F. Shahidi, In silico analysis of bioactive peptides produced from underutilized sea cucumber by-products—A bioinformatics approach, Mar. Drugs, 2022, 20, 1–16 CrossRef PubMed.
- F. Shahidi and A. Hossain, Preservation of aquatic food using edible films and coatings containing essential oils: A review, Crit. Rev. Food Sci. Nutr., 2022, 62, 66–105 CrossRef CAS PubMed.
- Y. Liu, N. Wang, Y. Tian, Y. Chang and J. Wang, Fucoidans from Thelenota ananas with 182.4 kDa Exhibited Optimal Anti-Adipogenic Activities by Modulating the Wnt/β-Catenin Pathway, J. Ocean Univ. China, 2021, 20, 921–930 CrossRef CAS.
- R. J. Fonseca and P. A. Mourão, Pharmacological activities of sulfated fucose-rich polysaccharides after oral administration: Perspectives for the development of new carbohydrate-based drugs, Mar. Drugs, 2021, 19, 1–20 CrossRef PubMed.
- G. Li, Y. Zhou, W. Y. Yang, C. Zhang, L. Hong and L. Jia, Inhibitory effects of sulfated polysaccharides from the sea cucumber Cucumaria frondosa against Aβ40 aggregation and cytotoxicity, ACS Chem. Neurosci., 2021, 12, 1854–1859 CrossRef CAS PubMed.
- D. Weedon, Marine injuries, in Weedon's Skin Pathology, 3rd edn, 2010, pp. 641–643 Search PubMed.
- M. A. Sibiya, J. Jeyavani, J. Sivakamavalli, C. Ravi, M. Divya and B. Vaseeharan, Bioactive compounds from various types of sea urchin and their therapeutic effects—a review, Reg. Stud. Mar. Sci., 2021, 44, 101760 Search PubMed.
- A. Hossain, D. Dave and F. Shahidi, Sulfated polysaccharides in sea cucumbers and their biological properties: A review, Int. J. Biol. Macromol., 2023, 253, 1–22 Search PubMed.
- J. Gao, L. Lin, B. Sun and M. Zhao, A comparison study on polysaccharides extracted from Laminaria japonica using different methods: Structural characterization and bile acid-binding capacity, Food Funct., 2017, 8, 3043–3052 RSC.
- Q. Zhu, L. Lin and M. Zhao, Sulfated fucan/fucosylated chondroitin sulfate-dominated polysaccharide fraction from low-edible-value sea cucumber ameliorates type 2 diabetes in rats: New prospects for sea cucumber polysaccharide based-hypoglycemic functional food, Int. J. Biol. Macromol., 2020, 159, 34–45 CrossRef CAS PubMed.
- Y. Gao, Z. Li, Y. Qi, Z. Guo, Y. Lin, W. Li, Y. Hu and Q. Zhao, Proximate composition and nutritional quality of deep sea growth sea cucumbers (Stichopus japonicus) from different origins, J. Sci. Food Agric., 2016, 96, 2378–2383 CrossRef CAS PubMed.
- M. P. Myron, S. Siddiquee and S. A. Azad, Partial structural studies of fucosylated chondroitin sulfate (FuCS) using attenuated total reflection fourier transform infrared spectroscopy (ATR-FTIR) and chemometrics, Vib. Spectrosc., 2017, 89, 26–36 CrossRef.
- Z. Zhu, B. Zhu, Y. Sun, C. Ai, L. Wang, C. Wen, J. Yang, S. Song and X. Liu, Sulfated polysaccharide from sea cucumber and its depolymerized derivative prevent obesity in association with modification of gut microbiota in high-fat diet-fed mice, Mol. Nutr. Food Res., 2018, 62, 1–12 Search PubMed.
- G. R. Santos, B. F. Glauser, L. A. Parreiras, E. Vilanova and P. A. Mourão, Distinct structures of the α-fucose branches in fucosylated chondroitin sulfates do not affect their anticoagulant activity, Glycobiology, 2015, 25, 1043–1052 CrossRef CAS PubMed.
- M. Wu, L. Xu, L. Zhao, C. Xiao, N. Gao, L. Luo, L. Yang, Z. Li, L. Chen and J. Zhao, Structural analysis and anticoagulant activities of the novel sulfated fucan possessing a regular well-defined repeating unit from sea cucumber, Mar. Drugs, 2015, 13, 2063–2084 CrossRef CAS PubMed.
- M. B. Mansour, R. Balti, L. Yacoubi, V. Ollivier, F. Chaubet and R. M. Maaroufi, Primary structure and anticoagulant activity of fucoidan from the sea cucumber Holothuria polii, Int. J. Biol. Macromol., 2019, 121, 1145–1153 CrossRef PubMed.
- Y. Zhu, Y. Tian, N. Wang, Y. Chang, C. Xue and J. Wang, Structure–function relationship analysis of fucoidan from sea cucumber (Holothuria tubulosa) on ameliorating metabolic inflammation, J. Food Biochem., 2021, 45, 1–10 Search PubMed.
- D. Shi, J. Qi, H. Zhang, H. Yang, Y. Yang and X. Zhao, Comparison of hydrothermal depolymerization and oligosaccharide profile of fucoidan and fucosylated chondroitin sulfate from Holothuria floridana, Int. J. Biol. Macromol., 2019, 132, 738–747 CrossRef CAS PubMed.
- Z. Wei and Y. Chang, The Extraction, Separation Technology, and, New Product Development of Sulfated Polysaccharides from Sea Cucumber, in Advances in Sea Cucumber Processing Technology and Product Development, ed. C. Xue, Springer, Cham, 2022, pp. 229–257 Search PubMed.
- Q. Xiong, Z. Song, W. Hu, J. Liang, Y. Jing, L. He and S. Li, Methods of extraction, separation, purification, structural characterization for polysaccharides from aquatic animals and their major pharmacological activities, Crit. Rev. Food Sci. Nutr., 2020, 60, 48–63 CrossRef CAS PubMed.
- M. I. Arnone, M. Byrne and P. Martinez, Echinodermata, in Evolutionary Developmental Biology of Invertebrates, ed. A. Wanninger, Springer, Vienna, 2015, pp. 1–58 Search PubMed.
- V. H. Pomin, Holothurian fucosylated chondroitin sulfates, Mar. Drugs, 2014, 12, 232–254 CrossRef PubMed.
- N. E. Ustyuzhanina, M. I. Bilan, A. S. Dmitrenok, E. A. Tsvetkova, A. S. Shashkov, V. A. Stonik, N. E. Nifantiev and A. I. Usov, Structural characterization of fucosylated chondroitin sulfates from sea cucumbers Apostichopus japonicus and Actinopyga mauritiana, Carbohydr. Polym., 2016, 153, 399–405 CrossRef CAS PubMed.
- P. X. Gong, Q. Y. Li, Y. C. Wu, W. Y. Lu, J. Zeng and H. J. Li, Structural elucidation and antidiabetic activity of fucosylated chondroitin sulfate from sea cucumber Stichopus japonicas, Carbohydr. Polym., 2021, 262, 1–9 CrossRef PubMed.
- N. E. Ustyuzhanina, M. I. Bilan, N. Y. Anisimova, A. S. Dmitrenok, E. A. Tsvetkova, M. V. Kiselevskiy, N. E. Nifantiev and A. I. Usov, Depolymerization of a fucosylated chondroitin sulfate from Cucumaria japonica: Structure and activity of the product, Carbohydr. Polym., 2022, 281, 1–8 CrossRef PubMed.
- V. H. Pomin, Fucanomics and galactanomics: Current status in drug discovery, mechanisms of action and role of the well-defined structures, Biochim. Biophys. Acta, Gen. Subj., 2012, 1820, 1971–1979 CrossRef CAS PubMed.
- F. C. Teixeira, E. O. Kozlowski, K. V. D. A. Micheli, A. C. E. Vilela-Silva, L. Borsig and M. S. Pavão, Sulfated fucans and a sulfated galactan from sea urchins as potent inhibitors of selectin-dependent hematogenous metastasis, Glycobiology, 2018, 28, 427–434 CrossRef CAS PubMed.
- N. E. Ustyuzhanina, M. I. Bilan, A. S. Dmitrenok, E. Y. Borodina, N. E. Nifantiev and A. I. Usov, A highly regular fucan sulfate from the sea cucumber Stichopus horrens, Carbohydr. Res., 2018, 456, 5–9 CrossRef CAS PubMed.
- F. Shang, R. Mou, Z. Zhang, N. Gao, L. Lin, Z. Li, M. Wu and J. Zhao, Structural analysis and anticoagulant activities of three highly regular fucan sulfates as novel intrinsic factor Xase inhibitors, Carbohydr. Polym., 2018, 195, 257–266 CrossRef CAS PubMed.
- V. H. Pomin and P. A. Mourão, Structure, biology, evolution, and medical importance of sulfated fucans and galactans, Glycobiology, 2008, 18, 1016–1027 CrossRef CAS PubMed.
- W. Zhang, J. Wang, W. Jin and Q. Zhang, The antioxidant activities and neuroprotective effect of polysaccharides from the starfish Asterias rollestoni, Carbohydr. Polym., 2013, 95, 9–15 CrossRef CAS PubMed.
- Y. Liu, X. Wu, Y. Wang, W. Jin and Y. Guo, The immunoenhancement effects of starfish Asterias rollestoni polysaccharides in macrophages and cyclophosphamide-induced immunosuppression mouse models, Food Funct., 2020, 11, 10700–10708 RSC.
- R. Ramachandra, R. B. Namburi, O. Ortega-Martinez, X. Shi, J. Zaia, S. T. Dupont, M. C. Thorndyke, U. Lindahl and D. Spillmann, Brittlestars contain highly sulfated chondroitin sulfates/dermatan sulfates that promote fibroblast growth factor 2-induced cell signaling, Glycobiology, 2014, 24, 195–207 CrossRef CAS PubMed.
- H. Ma, Q. Yuan, H. Tang, H. Tan, T. Li, S. Wei, J. Huang, Y. Yao, Y. Hu, S. Zhong, Y. Liu, C. Gao and L. Zhao, Structural elucidation of a glucan from Trichaster palmiferus by its degraded products and preparation of its sulfated derivative as an anticoagulant, Mar. Drugs, 2023, 21, 1–14 CrossRef PubMed.
- G. Chen, J. Shen, Y. Zhang, F. Shi, X. Mei, C. Xue and Y. Chang, Sulfated fucan could serve as a species marker of sea cucumber with endo-1, 3-fucanase as the essential tool, Carbohydr. Polym., 2023, 312, 1 CrossRef PubMed.
- A. Hossain, D. Dave and F. Shahidi, Northern sea cucumber (Cucumaria frondosa): A potential candidate for functional food, nutraceutical, and pharmaceutical sector, Mar. Drugs, 2020, 18, 1–27 CrossRef PubMed.
- T. Nakano, M. Betti and Z. Pietrasik, Extraction, isolation and analysis of chondroitin sulfate glycosaminoglycans, Recent Pat. Food, Nutr. Agric., 2010, 2, 61–74 CAS.
- J. Mou, Q. Li, X. Qi and J. Yang, Structural comparison, antioxidant and anti-inflammatory properties of fucosylated chondroitin sulfate of three edible sea cucumbers, Carbohydr. Polym., 2018, 185, 41–47 CrossRef CAS PubMed.
- L. Yang, Y. Wang, S. Yang and Z. Lv, Separation, purification, structures and anticoagulant activities of fucosylated chondroitin sulfates from Holothuria scabra, Int. J. Biol. Macromol., 2018, 108, 710–718 CrossRef CAS PubMed.
- R. Guan, Y. Peng, L. Zhou, W. Zheng, X. Liu, P. Wang, Q. Yuan, N. Gao, L. Zhao and J. Zhao, Precise structure and anticoagulant activity of fucosylated glycosaminoglycan from Apostichopus japonicus: Analysis of its depolymerized fragments, Mar. Drugs, 2019, 17, 1–13 CrossRef PubMed.
- L. Gao, C. Xu, X. Tao, Z. Zuo, Z. Ning, L. Wang, N. Gao and J. Zhao, Structure elucidation of fucan sulfate from sea cucumber Holothuria fuscopunctata through a bottom-up strategy and the antioxidant activity analysis, Int. J. Mol. Sci., 2022, 23, 1–18 CAS.
- N. E. Ustyuzhanina, M. I. Bilan, N. E. Nifantiev and A. I. Usov, Structural analysis of holothurian fucosylated chondroitin sulfates: Degradation versus non-destructive approach, Carbohydr. Res., 2019, 476, 8–11 CrossRef CAS PubMed.
- K. Wang, L. Qi, L. Zhao, J. Liu, Y. Guo and C. Zhang, Degradation of chondroitin sulfate: Mechanism of degradation, influence factors, structure-bioactivity relationship and application, Carbohydr. Polym., 2023, 301, 1–18 Search PubMed.
- M. P. Myron, S. Siddiquee and S. A. Al Azad, Fucosylated chondroitin sulfate diversity in sea cucumbers: A review, Carbohydr. Polym., 2014, 112, 173–178 CrossRef PubMed.
- V. H. Pomin, Sulfated glycans in sea urchin fertilization, Glycoconjugate J., 2015, 32, 9–15 CrossRef CAS PubMed.
- Y. Chang, Y. Hu and D. J. McClements, Competitive adsorption and displacement of anionic polysaccharides (fucoidan and gum arabic) on the surface of protein-coated lipid droplets, Food Hydrocolloids, 2016, 52, 820 CrossRef CAS.
- P. A. G. Soares, K. A. Ribeiro, A. P. Valente, N. V. Capille, S. N. M. C. G. Oliveira, A. M. F. Tovar, M. S. Pereira, E. Vilanova and P. A. S. Mourao, A unique fucosylated chondroitin sulfate type II with strikingly homogeneous and neatly distributed α-fucose branches, Glycobiology, 2018, 28, 565–579 CrossRef CAS PubMed.
- Q. Li, S. Jiang, W. Shi, X. Qi, W. Song, J. Mou and J. Yang, Structure characterization, antioxidant and immunoregulatory properties of a novel fucoidan from the sea cucumber Stichopus chloronotus, Carbohydr. Polym., 2020, 231, 1–9 Search PubMed.
- G. Chen, L. Yu, F. Shi, J. Shen, Y. Zhang, G. Liu, X. Mei, X. Li, X. Xu, C. Xue and Y. Chang, A comprehensive review of sulfated fucan from sea cucumber: Antecedent and prospect, Carbohydr. Polym., 2024, 341, 1–13 CrossRef PubMed.
- S. Chen, Y. Hu, X. Ye, G. Li, G. Yu, C. Xue and W. Chai, Sequence determination and anticoagulant and antithrombotic activities of a novel sulfated fucan isolated from the sea cucumber Isostichopus badionotus, Biochim. Biophys. Acta, Gen. Subj., 2012, 1820, 989–1000 CrossRef CAS PubMed.
- G. Chen, L. Yu, Y. Zhang, Y. Chang, Y. Liu, J. Shen and C. Xue, Utilizing heterologously overexpressed endo-1, 3-fucanase to investigate the structure of sulfated fucan from sea cucumber (Holothuria hilla), Carbohydr. Polym., 2021, 272, 1–6 Search PubMed.
- Y. Cai, W. Yang, R. Yin, L. Zhou, Z. Li, M. Wu and J. Zhao, An anticoagulant fucan sulfate with hexasaccharide repeating units from the sea cucumber Holothuria albiventer, Carbohydr. Res., 2018, 464, 12 CrossRef CAS PubMed.
- R. Dwivedi, M. Farrag, P. Sharma, D. Shi, A. A. Shami, S. K. Misra, P. Ray, J. Shukla, F. Zhang, R. J. Linhardt, J. S. Sharp, R. Tandon and V. H. Pomin, The sea cucumber Thyonella gemmata contains a low anticoagulant sulfated fucan with high anti-SARS-CoV-2 actions against wild-type and delta variants, J. Nat. Prod., 2023, 86, 1463–1475 CrossRef CAS PubMed.
- W. Zheng, L. Zhou, L. Lin, Y. Cai, H. Sun, L. Zhao, N. Gao, R. Yin and J. Zhao, Physicochemical characteristics and anticoagulant activities of the polysaccharides from sea cucumber Pattalus mollis, Mar. Drugs, 2019, 17, 1–15 CAS.
- P. D. Thinh, B. M. Ly, R. V. Usoltseva, N. M. Shevchenko, A. B. Rasin, S. D. Anastyuk, O. S. Malyarenko, T. N. Zvyagintseva, P. T. San and S. P. Ermakova, A novel sulfated fucan from Vietnamese sea cucumber Stichopus variegatus: Isolation, structure and anticancer activity in vitro, Int. J. Biol. Macromol., 2018, 117, 1101–1109 CrossRef CAS PubMed.
- N. Gao, R. Chen, R. Mou, J. Xiang, K. Zhou, Z. Li and J. Zhao, Purification, structural characterization and anticoagulant activities of four sulfated polysaccharides from sea cucumber Holothuria fuscopunctata, Int. J. Biol. Macromol., 2020, 164, 3421–3428 CrossRef CAS PubMed.
- N. E. Ustyuzhanina, M. I. Bilan, A. S. Dmitrenok, A. S. Shashkov, N. E. Nifantiev and A. I. Usov, The structure of a fucosylated chondroitin sulfate from the sea cucumber Cucumaria frondosa, Carbohydr. Polym., 2017, 165, 7–12 CrossRef CAS PubMed.
- J. Yang, Y. Wang, T. Jiang and Z. Lv, Novel branch patterns and anticoagulant activity of glycosaminoglycan from sea cucumber Apostichopus japonicus, Int. J. Biol. Macromol., 2015, 72, 911–918 CrossRef CAS PubMed.
- H. Xu, Q. Zhou, B. Liu, F. Chen and M. Wang, Holothurian fucosylated chondroitin sulfates and their potential benefits for human health: Structures and biological activities, Carbohydr. Polym., 2022, 275, 118691 CrossRef CAS PubMed.
- Y. G. Shi, Y. C. Meng, J. R. Li, J. Chen, Y. H. Liu and X. Bai, Chondroitin sulfate: Extraction, purification, microbial and chemical synthesis, J. Chem. Technol. Biotechnol., 2014, 89, 1445–1465 CrossRef CAS.
- N. E. Ustyuzhanina, M. I. Bilan, A. S. Dmitrenok, A. S. Shashkov, N. M. A. Ponce, C. A. Stortz, N. E. Nifantiev and A. I. Usov, Fucosylated chondroitin sulfate from the sea cucumber Hemioedema spectabilis: Structure and influence on cell adhesion and tubulogenesis, Carbohydr. Polym., 2020, 234, 1–8 CrossRef PubMed.
- X. Liu, Y. Liu, J. Hao, X. Zhao, Y. Lang, F. Fan, C. Cai, G. Li, L. Zhang and G. Yu, In vivo anti-cancer mechanism of low-molecular-weight fucosylated chondroitin sulfate (LFCS) from sea cucumber Cucumaria frondosa, Molecules, 2016, 21, 1–12 CrossRef PubMed.
- M. B. Mansour, R. Balti, V. Ollivier, H. B. Jannet, F. Chaubet and R. M. Maaroufi, Characterization and anticoagulant activity of a fucosylated chondroitin sulfate with unusually procoagulant effect from sea cucumber, Carbohydr. Polym., 2017, 174, 760–771 CrossRef PubMed.
- Z. Zhu, X. Dong, C. Yan, C. Ai, D. Zhou, J. Yang, H. Zhang, X. Liu, S. Song, H. Xiao and B. Zhu, Structural features and digestive behavior of fucosylated chondroitin sulfate from sea cucumbers Stichopus japonicus, J. Agric. Food Chem., 2019, 67, 10534–10542 CrossRef CAS PubMed.
- Q. Li, C. Cai, Y. Chang, F. Zhang, R. J. Linhardt, C. Xue, G. Li and G. Yu, A novel structural fucosylated chondroitin sulfate from Holothuria mexicana and its effects on growth factors binding and anticoagulation, Carbohydr. Polym., 2018, 181, 1160–1168 CrossRef CAS PubMed.
- S. Chen, C. Xue, Q. Tang, G. Yu and W. Chai, Comparison of structures and anticoagulant activities of fucosylated chondroitin sulfates from different sea cucumbers, Carbohydr. Polym., 2011, 83, 688–696 CrossRef CAS.
- M. O. Castro, V. H. Pomin, L. L. Santos, A.-C. E. S. Vilela-Silva, N. Hirohashi, L. Pol-Fachin, H. Verli and P. A. S. Mourão, A unique 2-sulfated β-galactan from the egg jelly of the sea urchin Glyptocidaris crenularis, J. Biol. Chem., 2009, 284, 18790–18800 CrossRef CAS PubMed.
- N. E. Ustyuzhanina, M. I. Bilan, A. S. Dmitrenok, E. A. Tsvetkova, N. E. Nifantiev and A. I. Usov, Oversulfated dermatan sulfate and heparinoid in the starfish Lysastrosoma anthosticta: Structures and anticoagulant activity, Carbohydr. Polym., 2021, 261, 1–9 CrossRef PubMed.
- R. Li, H. Yu, Y. Yue, S. Liu, R. E. Xing, X. Chen and P. Li, Sulfated polysaccharides with antioxidant and anticoagulant activity from the sea cucumber Holothuria fuscogliva, Chin. J. Oceanol. Limnol., 2017, 35, 763–769 CrossRef CAS.
- N. Wu, Y. Zhang, X. Ye, Y. Hu, T. Ding and S. Chen, Sulfation pattern of fucose branches affects the anti-hyperlipidemic activities of fucosylated chondroitin sulfate, Carbohydr. Polym., 2016, 147, 1–7 CrossRef CAS PubMed.
- H. Jiao, X. Shang, Q. Dong, S. Wang, X. Liu, H. Zheng and X. Lu, Polysaccharide constituents of three types of sea urchin shells and their anti-inflammatory activities, Mar. Drugs, 2015, 13, 5882–5900 CrossRef CAS PubMed.
- J. Mou, D. Zhuang, Q. Li, W. Song and J. Yang, Comparison of chain conformation properties of bio-active fucosylated chondroitin sulfates from two different sea cucumbers, Int. J. Biol. Macromol., 2019, 133, 44–50 CrossRef CAS PubMed.
- C. G. Panagos, D. S. Thomson, C. Moss, A. D. Hughes, M. S. Kelly, Y. Liu, W. Chai, R. Venkatasamy, D. Spina, C. P. Page, J. P. Hogwood, R. J. Woods, B. Mulloy, C. D. Bavington and D. Uhrín, Fucosylated chondroitin sulfates from the body wall of the sea cucumber Holothuria forskali: Conformation, selectin binding, and biological activity, J. Biol. Chem., 2014, 289, 28284–28298 CrossRef CAS PubMed.
- X. Dong, R. Pan, X. Deng, Y. Chen, G. Zhao and C. Wang, Separation, purification, anticoagulant activity and preliminary structural characterization of two sulfated polysaccharides from sea cucumber Acaudina molpadioidea and Holothuria nobilis, Process Biochem., 2014, 49, 1352–1361 CrossRef CAS.
- M. Wu, R. Huang, D. Wen, N. Gao, J. He, Z. Li and J. Zhao, Structure and effect of sulfated fucose branches on anticoagulant activity of the fucosylated chondroitin sulfate from sea cucumber Thelenata ananas, Carbohydr. Polym., 2012, 87, 862–868 CrossRef CAS PubMed.
- H. Ghelani, M. Khursheed, T. E. Adrian and R. K. Jan, Anti-inflammatory effects of compounds from echinoderms, Mar. Drugs, 2022, 20, 1–24 CrossRef PubMed.
- Y. Chen, Y. Wang, S. Yang, M. Yu, T. Jiang and Z. Lv, Glycosaminoglycan from Apostichopus japonicus improves glucose metabolism in the liver of insulin resistant mice, Mar. Drugs, 2019, 18, 1–14 CrossRef CAS PubMed.
- S. Li, W. Jiang, S. Hu, W. Song, L. Ji, Y. Wang and L. Cai, Fucosylated chondroitin sulphate from Cucumaria frondosa mitigates hepatic endoplasmic reticulum stress and inflammation in insulin resistant mice, Food Funct., 2015, 6, 1547–1556 RSC.
- M. M. Stefaniak-Vidarsson, V. A. Kale, M. Gudjónsdóttir, G. Marteinsdottir, O. Fridjonsson, G. O. Hreggvidsson, O. E. Sigurjonsson, S. Omarsdottir and K. Kristbergsson, Bioactive effect of sulphated polysaccharides derived from orange-footed sea cucumber (Cucumaria frondosa) toward THP-1 macrophages, Bioact. Carbohydr. Diet. Fibre, 2017, 12, 14–19 CrossRef CAS.
- L. Olivera-Castillo, G. Grant, N. Kantún-Moreno, H. A. Barrera-Pérez, J. Montero, M. A. Olvera-Novoa, L. M. Carrillo-Cocom, J. J. Acevedo, C. Puerto-Castillo, V. M. Solís and J. A. Pérez-Vega, A glycosaminoglycan-rich fraction from sea cucumber Isostichopus badionotus has potent anti-inflammatory properties in vitro and in vivo, Nutrients, 2020, 12, 1–15 CrossRef PubMed.
- J. Wang, S. Hu, W. Jiang, W. Song, L. Cai and J. Wang, Fucoidan from sea cucumber may improve hepatic inflammatory response and insulin resistance in mice, Int. Immunopharmacol., 2016, 31, 15–23 CrossRef CAS PubMed.
- J. Yin, X. Yang, B. Xia, Z. Yang, Z. Wang, J. Wang, T. Li, P. Lin, X. Song and S. Guo, The fucoidan from sea cucumber Apostichopus japonicus attenuates lipopolysaccharide-challenged liver injury in C57BL/6J mice, J. Funct. Foods, 2019, 61, 1–13 Search PubMed.
- X. Xu, Y. Chang, C. Xue, J. Wang and J. Shen, Gastric protective activities of sea cucumber fucoidans with different molecular weight and chain conformations: a structure–activity relationship investigation, J. Agric. Food Chem., 2018, 66, 8615–8622 CrossRef CAS PubMed.
- J. Li, S. Li, L. Wu, H. Yang, C. Wei, T. Ding, R. J. Linhardt, X. Zheng, X. Ye and S. Chen, Ultrasound-assisted fast preparation of low molecular weight fucosylated chondroitin sulfate with antitumor activity, Carbohydr. Polym., 2019, 209, 82–91 CrossRef CAS PubMed.
- Y. B. Salem, S. Amri, K. M. Hammi, A. Abdelhamid, D. Le Cerf, A. Bouraoui and H. Majdoub, Physico-chemical characterization and pharmacological activities of sulfated polysaccharide from sea urchin, Paracentrotus lividus, Int. J. Biol. Macromol., 2017, 97, 8–15 CrossRef PubMed.
- F. Shahidi and A. Saeid, Bioactivity of marine-derived peptides and proteins: a review, Mar. Drugs, 2025, 23, 157 CrossRef CAS PubMed.
- S. C. Lourenço, M. Moldão-Martins and V. D. Alves, Antioxidants of natural plant origins: From sources to food industry applications, Molecules, 2019, 24, 1–25 CrossRef PubMed.
- L. Yu, C. Xue, Y. Chang, X. Xu, L. Ge, G. Liu and Y. Wang, Structure elucidation of fucoidan composed of a novel tetrafucose repeating unit from sea cucumber, Thelenota ananas, Food Chem., 2014, 146, 113–119 CrossRef CAS PubMed.
- Q. Zhong, B. Wei, S. Wang, S. Ke, J. Chen, H. Zhang and H. Wang, The antioxidant activity of polysaccharides derived from marine organisms: An overview, Mar. Drugs, 2019, 17, 1–34 CrossRef PubMed.
- J. Mou, C. Wang, Q. Li, X. Qi and J. Yang, Preparation and antioxidant properties of low molecular holothurian glycosaminoglycans by H2O2/ascorbic acid degradation, Int. J. Biol. Macromol., 2018, 107, 1339–1347 CrossRef CAS PubMed.
- H. Yin, R. Li, J. Liu, Y. Sun, L. Zhao, J. Mou and J. Yang, Fucosylated chondroitin sulfate from sea cucumber Stichopus chloronotus alleviate the intestinal barrier injury and oxidative stress damage in vitro and in vivo, Carbohydr. Polym., 2024, 328, 1–12 CrossRef PubMed.
- J. Wang, S. Shi, F. Li, X. Du, B. Kong, H. Wang and X. Xia, Physicochemical properties and antioxidant activity of polysaccharides obtained from sea cucumber gonads via ultrasound-assisted enzymatic techniques, LWT – Food Sci. Technol., 2022, 160, 1–9 Search PubMed.
- H. K. Matthews, C. Bertoli and R. A. M. de Bruin, Cell cycle control in cancer, Nat. Rev. Mol. Cell Biol., 2022, 23, 74–88 CrossRef CAS PubMed.
- K. K. Sanjeewa and K. H. I. N. M. Herath, Bioactive secondary metabolites in sea cucumbers and their potential to use in the functional food industry, Fish. Aquat. Sci., 2023, 26, 69–86 CrossRef.
- J. L. Kim, S. H. Park, S. Jeong, B. R. Kim, Y. J. Na, M. J. Jo, Y. A. Jeong, H. K. Yun, D. Y. Kim, B. G. Kim, S. You, S. C. Oh and D. H. Lee, Sea cucumber (Stichopus japonicas) F2 enhanced TRAIL-induced apoptosis via XIAP ubiquitination and ER stress in colorectal cancer cells, Nutrients, 2019, 11, 1–14 Search PubMed.
- M. Zhang, L. Chen, Y. Liu, M. Chen, S. Zhang and D. Kong, Sea cucumber Cucumaria frondosa fucoidan inhibits osteosarcoma adhesion and migration by regulating cytoskeleton remodeling, Oncol. Rep., 2020, 44, 469–476 CrossRef CAS PubMed.
- U. Surayot, S. Lee and S. You, Effects of sulfated fucan from the sea cucumber Stichopus japonicus on natural killer cell activation and cytotoxicity, Int. J. Biol. Macromol., 2018, 108, 177–184 CrossRef CAS PubMed.
- H. Wang, S. Yang, Y. Wang, T. Jiang, S. Li and Z. Lv, Immunoenhancement effects of glycosaminoglycan from Apostichopus japonicus: In vitro and in cyclophosphamide-induced immunosuppressed mice studies, Mar. Drugs, 2017, 15, 1–14 Search PubMed.
- C. Lin, X. Zhu, Q. Jin, A. Sui, J. Li and L. Shen, Effects of holothurian glycosaminoglycan on the sensitivity of lung cancer to chemotherapy, Integr. Cancer Ther., 2020, 19, 1–10 Search PubMed.
- C. Liu, Q. Lin, Y. Gao, L. Ye, Y. Xing and T. Xi, Characterization and antitumor activity of a polysaccharide from Strongylocentrotus nudus eggs, Carbohydr. Polym., 2007, 67(3), 313–318 CrossRef CAS.
- X. H. Shang, X. Y. Liu, J. P. Zhang, Y. Gao, B. H. Jiao, H. Zheng and X. L. Lu, Traditional Chinese Medicine—Sea Urchin, Mini-Rev. Med. Chem., 2014, 14, 537–542 CrossRef CAS PubMed.
- Z. L. Zhang, C. Zhang, H. Liao, Y. M. Liu, W. N. Cheng and J. Y. Du, Antitumor activities of extractive from sea urchin in vitro, Chin. J. Cancer Prev. Treat., 2003, 10, 569–572 Search PubMed.
- K. S. Kyung-Soo, Breast cancer chemopreventive activity of polysaccharides from starfish in vitro, J. Microbiol. Biotechnol., 2006, 16, 1405–1409 Search PubMed.
- K. S. Lee, J. S. Shin and K. S. Nam, Starfish polysaccharides downregulate metastatic activity through the MAPK signaling pathway in MCF-7 human breast cancer cells, Mol. Biol. Rep., 2013, 40, 5959–5966 CrossRef CAS PubMed.
- K. S. Lee, J. S. Shin and K. S. Nam, Cancer Chemopreventive Effects of Starfish Polysaccharide in Human Breast Cancer Cells, Biotechnol. Bioprocess Eng., 2011, 16, 987–991 CrossRef CAS.
- K. S. Nam and Y. H. Shon, Chemopreventive effects of polysaccharides extract from Asterina pectinifera on HT-29 human colon adenocarcinoma cells, BMB Rep., 2009, 42, 277–280 CrossRef CAS PubMed.
- J. Baharara and E. Amini, The potential of brittle star extracted polysaccharide in promoting apoptosis via intrinsic signaling pathway, Avicenna J. Med. Biotechnol., 2015, 7, 151–158 Search PubMed.
- J. Baharara, E. Amini and M. Musavi, Anti-vasculogenic activity of a polysaccharide derived from brittle star via inhibition of VEGF, paxillin and MMP-9, Iran. J. Biotechnol., 2017, 15, 179–185 CrossRef PubMed.
- L. Gozzo, P. Viale, L. Longo, D. C. Vitale and F. Drago, The potential role of heparin in patients with COVID-19: beyond the anticoagulant effect. A review, Front. Pharmacol., 2020, 11, 1–8 CrossRef PubMed.
- R. Yin, Y. Pan, Y. Cai, F. Yang, N. Gao, D. Ruzemaimaiti and J. Zhao, Re-understanding of structure and anticoagulation: Fucosylated chondroitin sulfate from sea cucumber Ludwigothurea grisea, Carbohydr. Polym., 2022, 294, 1–12 CrossRef PubMed.
- Y. Liu, R. Li, L. Song, K. Li, H. Yu, R. Xing, S. Liu and P. Li, Intermediate molecular weight-fucosylated chondroitin sulfate from sea cucumber Cucumaria frondosa is a promising anticoagulant targeting intrinsic factor IXa, Int. J. Biol. Macromol., 2024, 269, 1–10 Search PubMed.
- Q. Yuan, H. Li, Q. Wang, S. Sun, Z. Fang, H. Tang, X. Shi, J. Wen, L. Huang, M. Bai, S. Zhong, Y. Liu, C. Gao and L. Zhao, Deaminative-cleaved, S. monotuberculatus fucosylated glycosaminoglycan: Structural elucidation and anticoagulant activity, Carbohydr. Polym., 2022, 298, 1–11 CrossRef PubMed.
- L. Chahed, R. Balti, S. Elhiss, N. Bouchemal, N. Ajzenberg, V. Ollivier, F. Chaubet, R. M. Maaroufi and M. B. Mansour, Anticoagulant activity of fucosylated chondroitin sulfate isolated from Cucumaria syracusana, Process Biochem., 2020, 91, 149 CrossRef CAS.
- L. Cui, H. Sun, X. Shang, J. Wen, P. Li, S. Yang, L. Chen, X. Huang, H. Li, R. Yin and J. Zhao, Purification and Structural Analyses of Sulfated Polysaccharides from Low-Value Sea Cucumber Stichopus naso and Anticoagulant Activities of Its Oligosaccharides, Mar. Drugs, 2024, 22(265), 1–15 Search PubMed.
- P. Tian, D. Zhou, C. Ji, C. Niu, Y. Chen and Y. Chen, Characterization and anticoagulant activity of a fucosylated chondroitin sulfate from the sea cucumber Acaudina leucoprocta, Process Biochem., 2024, 147, 130–136 CrossRef CAS.
- W. He, H. Sun, L. Su, D. Zhou, X. Zhang, S. Shanggui and Y. Chen, Structure and anticoagulant activity of a sulfated fucan from the sea cucumber Acaudina leucoprocta, Int. J. Biol. Macromol., 2020, 164, 87–94 CrossRef CAS PubMed.
- Y. Ma, N. Gao, Z. Zuo, S. Li, W. Zheng, X. Shi, Q. Liu, T. Ma, R. Yin, X. Li and J. Zhao, Five distinct fucan sulfates from sea cucumber Pattalus mollis: Purification, structural characterization and anticoagulant activities, Int. J. Biol. Macromol., 2021, 186, 535–543 CrossRef CAS PubMed.
- Z. Ning, P. Wang, Z. Zuo, X. Tao, L. Gao, C. Xu, Z. Wang, B. Wu, N. Gao and J. Zhao, A fucan sulfate with pentasaccharide repeating units from the sea cucumber Holothuria floridana and its anticoagulant activity, Mar. Drugs, 2022, 20, 1–19 Search PubMed.
- M. S. Pereira, A. C. E. Vilela-Silva, A. P. Valente and P. A. Mourão, A 2-sulfated, 3-linked α-L-galactan is an anticoagulant polysaccharide, Carbohydr. Res., 2002, 337, 2231–2238 CrossRef CAS PubMed.
- A. Sibiya, J. Jeyavani, J. Sivakamavalli, C. Ravi, M. Divya and B. Vaseeharan, Bioactive compounds from various types of sea urchin and their therapeutic effects—a review, Reg. Stud. Mar. Sci., 2021, 44, 1–8 Search PubMed.
- A. A. Vasconcelos, I. D. Sucupira, A. L. Guedes, I. N. Queiroz, F. S. Frattani, R. J. Fonseca and V. H. Pomin, Anticoagulant and antithrombotic properties of three structurally correlated sea urchin sulfated glycans and their low-molecular-weight derivatives, Mar. Drugs, 2018, 16, 304 CrossRef PubMed.
- K. Kc, S. Shakya and H. Zhang, Gestational diabetes mellitus and macrosomia: a literature review, Ann. Nutr. Metab., 2015, 66, 14–20 CrossRef CAS PubMed.
- J. S. Bhatti, A. Sehrawat, J. Mishra, I. S. Sidhu, U. Navik, N. Khullar, S. Kumar, G. K. Bhatti and P. H. Reddy, Oxidative stress in the pathophysiology of type 2 diabetes and related complications: Current therapeutics strategies and future perspectives, Free Radicals Biol. Med., 2022, 184, 114 CrossRef CAS PubMed.
- F. Zhao, Q. Liu, J. Cao, Y. Xu, Z. Pei, H. Fan, Y. Yuan, X. Shen and C. Li, A sea cucumber (Holothuria leucospilota) polysaccharide improves the gut microbiome to alleviate the symptoms of type 2 diabetes mellitus in Goto-Kakizaki rats, Food Chem. Toxicol., 2020, 135, 1–9 CrossRef PubMed.
- S. Hu, J. Wang, H. Xu, Y. Wang, Z. Li and C. Xue, Fucosylated chondroitin sulphate from sea cucumber inhibits high-fat-sucrose diet-induced apoptosis in mouse pancreatic islets via down-regulating mitochondrial signaling pathway, J. Funct. Foods, 2014, 7, 517–526 CrossRef CAS.
- Y. Wang, J. Wang, Y. Zhao, S. Hu, D. Shi and C. Xue, Fucoidan from sea cucumber Cucumaria frondosa exhibits anti-hyperglycemic effects in insulin resistant mice via activating the PI3K/PKB pathway and GLUT4, J. Biosci. Bioeng., 2016, 121, 36–42 CrossRef CAS PubMed.
- S. Hu, J. Wang, Y. Xu, H. Yang, J. Wang, C. Xue, X. Yan and L. Su, Anti-inflammation effects of fucosylated chondroitin sulphate from Acaudina molpadioides by altering gut microbiota in obese mice, Food Funct., 2019, 10, 1736–1746 RSC.
- S. Hu, S. Chen, H. Zhu, M. Du, W. Jiang, Y. Liu, X. Gao, L. Su and Y. Xu, Low molecular weight, 4-O-sulfation, and sulfation at meta-fucose positively promote the activities of sea cucumber fucoidans on improving insulin resistance in HFD-fed mice, Mar. Drugs, 2021, 20, 1–16 CrossRef PubMed.
- H. Xu, J. Wang, Y. Chang, J. Xu, Y. Wang, T. Long and C. Xue, Fucoidan from the sea cucumber Acaudina molpadioides exhibits anti-adipogenic activity by modulating the Wnt/β-catenin pathway and down-regulating the SREBP-1c expression, Food Funct., 2014, 5, 1547–1555 RSC.
- N. Huang, M. Y. Wu, C. B. Zheng, L. Zhu, J. H. Zhao and Y. T. Zheng, The depolymerized fucosylated chondroitin sulfate from sea cucumber potently inhibits HIV replication via interfering with virus entry, Carbohydr. Res., 2013, 380, 64–69 CrossRef CAS PubMed.
- S. Song, H. Peng, Q. Wang, Z. Liu, X. Dong, C. Wen, C. Ai, Y. Zhang, Z. Wang and B. Zhu, Inhibitory activities of marine sulfated polysaccharides against SARS-CoV-2, Food Funct., 2020, 11, 7415–7420 RSC.
- M. Ke, H. Wang, M. Zhang, Y. Tian, Y. Wang, B. Li, J. Yu, J. Dou, T. Xi and C. Zhou, The anti-lung cancer activity of SEP is mediated by the activation and cytotoxicity of NK cells via TLR2/4 in vivo, Biochem. Pharmacol., 2014, 89, 119–130 CrossRef CAS PubMed.
- R. Chen, W. Wang, R. Yin, Y. Pan, C. Xu, N. Gao, X. Luo and J. Zhao, Structural characterization and anticoagulant activities of a keratan sulfate-like polysaccharide from the sea cucumber Holothuria fuscopunctata, Mar. Drugs, 2023, 21, 632 CrossRef CAS PubMed.
- N. E. Ustyuzhanina, M. I. Bilan, A. S. Dmitrenok, A. S. Shashkov, M. I. Kusaykin, V. A. Stonik, N. E. Nifantiev and A. I. Usov, Structure and biological activity of a fucosylated chondroitin sulfate from the sea cucumber Cucumaria japonica, Glycobiology, 2016, 26, 449–459 CrossRef CAS PubMed.
- M. Zoepfl, R. Dwivedi, M. C. Taylor, V. H. Pomin and M. A. McVoy, Antiviral activities of four marine sulfated glycans against adenovirus and human cytomegalovirus, Antiviral Res., 2021, 190, 1–23 CrossRef PubMed.
- E. Garcia-Candela, C. Pariona-Velarde, A. Mondragón-Martínez and V. Chumpitaz-Cerrate, Antiviral activity of the sea cucumber tegument extract (Pattalus mollis) on human rotavirus A (RVA), Nat. Prod. Res., 2021, 35, 1014–1018 CrossRef CAS PubMed.
- S. Zou, R. Pan, X. Dong, M. He and C. Wang, Physicochemical properties and antioxidant activities of two fucosylated chondroitin sulfate from sea cucumber Acaudina molpadioidea and Holothuria nobilis, Process Biochem., 2016, 51, 650–658 CrossRef CAS.
- N. E. Wedamulla and W. A. J. P. Wijesinghe, Application of Polysaccharides in Food Technology: A Review, Trends Carbohydr. Res., 2021, 13, 35–49 CAS.
- L. E. Rioux, L. Beaulieu and S. L. Turgeon, Seaweeds: A traditional ingredient for a new gastronomic sensation, Food Hydrocolloids, 2017, 68, 255–265 CrossRef CAS.
- E. O. Mensah, O. N. Kanwugu, P. K. Panda and P. Adadi, Marine fucoidans: Structural, extraction, biological activities and their applications in the food industry, Food Hydrocolloids, 2023, 142, 1–25 CrossRef.
- O. Abdelhedi, A. Salem, R. Nasri, M. Nasri and M. Jridi, Food applications of bioactive marine gelatin films, Curr. Opin. Food Sci., 2022, 43, 206 CrossRef CAS.
- H. A. R. Suleria, S. Osborne, P. Masci and G. Gobe, Marine-based nutraceuticals: an innovative trend in the food and supplement industries, Mar. Drugs, 2015, 13, 6336–6351 CrossRef CAS PubMed.
- S. Ghosh, T. Sarkar, S. Pati, Z. A. Kari, H. A. Edinur and R. Chakraborty, Novel bioactive compounds from marine sources as a tool for functional food development, Front. Mar. Sci., 2022, 9, 1–28 Search PubMed.
- A. C. Freitas, L. Pereira, D. Rodrigues, A. P. Carvalho, T. Panteleitchouk, A. M. Gomes and A. C. Duarte, Marine Functional Foods, in Springer Handbook of Marine Biotechnology, ed. S. K. Kim, Springer Handbooks, Springer, Berlin, 2015, pp. 969–994 Search PubMed.
- L. Luo, M. Wu, L. Xu, W. Lian, J. Xiang, F. Lu, N. Gao, C. Xiao, S. Wang and J. Zhao, Comparison of physicochemical characteristics and anticoagulant activities of polysaccharides from three sea cucumbers, Mar. Drugs, 2013, 11, 399–417 CrossRef CAS PubMed.
- C. L. Okolie, B. Mason, A. Mohan, N. Pitts and C. C. Udenigwe, The comparative influence of novel extraction technologies on in vitro prebiotic-inducing chemical properties of fucoidan extracts from Ascophyllum nodosum, Food Hydrocolloids, 2019, 90, 462–471 CrossRef CAS.
- Y. Yuan and D. Macquarrie, Microwave assisted extraction of sulfated polysaccharides (fucoidan) from Ascophyllum nodosum and its antioxidant activity, Carbohydr. Polym., 2015, 129, 101–107 CrossRef CAS PubMed.
- A. F. Hifney, M. A. Fawzy, K. M. Abdel-Gawad and M. Gomaa, Industrial optimization of fucoidan extraction from Sargassum sp. and its potential antioxidant and emulsifying activities, Food Hydrocolloids, 2016, 54, 77–88 CrossRef CAS.
- L. Zhang, Q. Zhang, Y. Zheng, Z. He, P. Guan, X. He, L. Hui and Y. Dai, Study of Schiff base formation between dialdehyde cellulose and proteins, and its application for the deproteinization of crude polysaccharide extracts, Ind. Crops Prod., 2018, 112, 532–540 CrossRef CAS.
- R. Ghanbari, M. Zarei, A. Ebrahimpour, A. Abdul-Hamid, A. Ismail and N. Saari, Angiotensin-I converting enzyme (ACE) inhibitory and anti-oxidant activities of sea cucumber (Actinopyga lecanora) hydrolysates, Int. J. Mol. Sci., 2015, 16, 28870–28885 CrossRef CAS PubMed.
- Y. Zhang, S. Song, D. Song, H. Liang, W. Wang and A. Ji, Proliferative effects on neural stem/progenitor cells of a sulfated polysaccharide purified from the sea cucumber Stichopus japonicus, J. Biosci. Bioeng., 2010, 109, 67–72 CrossRef CAS PubMed.
- J. M. Heather and B. Chain, The sequence of sequencers: The history of sequencing DNA, Genomics, 2016, 107, 1–8 CrossRef CAS PubMed.
- R. Vitorino, S. Guedes, F. Trindade, I. Correia, G. Moura, P. Carvalho, M. A. S. Santos and F. Amado, De novo sequencing of proteins by mass spectrometry, Expert Rev. Proteomics, 2020, 17, 595–607 CrossRef CAS PubMed.
- Grand View Research, Chondroitin Sulfate Market Size, Share & Trends Analysis Report by Application (Nutraceuticals, Pharmaceuticals, and Cosmetics), by Region, and Segment Forecasts, 2024–2030, retrieved from: https://www.grandviewresearch.com/industry-analysis/chondroitin-sulfate-market, accessed on February 1, 2025.
- Acumen Research and Consulting, Chondroitin Sulfate Market Size, Share, and Trends Analysis Report, Acumen Research and Consulting, 2023, available at: https://www.acumenresearchandconsulting.com/press-releases/chondroitin-sulfate-market, accessed on February 1, 2025.
- T. Hou, Z. Wang, K. Tang, S. Zhang, S. Liu, J. Liu, X. Fan and X. Liu, The Fin-Improving Effects of Fucosylated Chondroitin Sulfate from Green and Purple Apostichopus japonicus on Caudal Fin Regeneration of Zebrafish Larvae, Aquacult. Res., 2023, 1–11 Search PubMed.
- W. Wen, L. Yang, X. Wang, H. Zhang, F. Wu, K. Xu, S. Chen and Z. Liao, Fucoidan promotes angiogenesis and accelerates wound healing through AKT/Nrf2/HIF-1α signalling pathway, Int. Wound J., 2023, 20, 3606–3618 CrossRef PubMed.
Footnote |
| † On leave of absence from the Department of Nutrition and Food Engineering, Daffodil International University, Dhaka-1216, Bangladesh. |
| This journal is © The Royal Society of Chemistry 2025 |