
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceEffect of seawater acidification on energy metabolism in the hydrocoral Millepora alcicornis: inhibition of citrate synthase activity indicates disruption in aerobic pathways†
Andrea Carlina
Jesulich
a,
Mariana Machado
Lauer
b,
Laura Fernandes de Barros
Marangoni
ac,
Joseane Aparecida
Marques
ac,
Yuri Dornelles
Zebral
b,
Cristiano Macedo
Pereira
c,
Gustavo Adolpho Santos
Duarte
d,
Débora de Oliveira
Pires
cd,
Clovis Barreira e
Castro
cd,
Emiliano Nicolas
Calderon
c and
Adalto
Bianchini
 *bc
*bc
aPós-Graduação Em Oceanografia Biológica, Instituto de Oceanografia, Universidade Federal Do Rio Grande, Av. Itália km 8, Rio Grande, RS 96203-900, Brazil
bInstituto de Ciências Biológicas, Universidade Federal Do Rio Grande, Av. Itália km 8, Campus Carreiros, Rio Grande, RS 96203-900, Brazil. E-mail: adaltobianchini@furg.br; Tel: +55 53 3293-5193
cInstituto Coral Vivo, Rua Dos Coqueiros 87, Santa Cruz Cabrália, BA 45807-000, Brazil
dMuseu Nacional, Universidade Federal do Rio de Janeiro, Parque Quinta da Boa Vista, São Cristovão, Rio de Janeiro, RJ 20940-040, Brazil
First published on 3rd February 2025
Abstract
Ocean acidification is a major threat to coral reefs worldwide, with reduced growth already reported in the hydrocoral Millepora alcicornis (Linnaeus, 1758) under these conditions. Inhibition of enzymes related to energy metabolism is hypothesized as one of the mechanisms associated with the physiological impacts of ocean acidification. Therefore, a mesocosm experiment was conducted to investigate whether three levels of decreasing seawater pH could alter the activity of key enzymes involved in the energy metabolism in M. alcicornis. Hydrocorals were acclimated to marine mesocosm conditions for 20 days and then exposed to different seawater pH levels [ambient pH (8.1) and experimental pH (7.8, 7.5 and 7.2)] for 16 and 30 days. Endpoints analyzed included the activity of key enzymes involved in the regulation of the glycolytic pathway (hexokinase and pyruvate kinase), aerobic energy production via the Krebs cycle (citrate synthase) and anaerobic energy production via lactate formation (lactate dehydrogenase). The results obtained show that only citrate synthase was affected by seawater acidification, as a marked reduction in its activity was observed at all experimental pH levels tested (7.8, 7.5 and 7.2). This finding indicates that reduced growth previously reported for M. alcicornis under seawater acidification conditions can be explained, at least in part, by a negative impact on the Krebs cycle, a major pathway involved in aerobic energy production.
Environmental significanceOcean acidification is a key major threat to aquatic biota, including coral reefs. Despite the recognized negative effects of this global environmental stressor on marine biota, little is known about its mechanism of action in corals and hydrocorals. In the present study, we evaluated seawater acidification effects on the activity of energy metabolism-related enzymes in the hydrocoral Millepora alcicornis. Our findings indicate that reduced growth previously reported for this hydrocoral species under seawater acidification conditions can be explained by a negative impact on the Krebs cycle, a major pathway involved in aerobic energy production. Negative impacts on the growth of reef calcifying organisms result in reduced ecological complexity and biodiversity of the entire coral reef system. |
Introduction
Ocean acidification refers to the decrease in seawater pH resulting from oceanic absorption of atmospheric CO2 emissions associated with anthropogenic sources. It leads to changes in both the carbonate buffer system and the calcium carbonate (CaCO3) saturation state of oceans. Indeed, the pH value of global seawater has remained stable at an average of 8.2 for millions of years. However, since industrialization began around the 1760s, ocean acidification has increased by 26% compared to the pre-industrial times and is expected to continue rising in the future. Indeed, the ambient pH of surface seawater was already reported to be 8.10.1 According to the latest Assessment Report (AR6) of the Intergovernmental Panel on Climate Change, it is projected to further drop by 0.3 to 0.4 units by 2100.2 This change would be equivalent to a 150% increase in seawater acidity compared to preindustrial times.Ocean acidification will likely affect all marine ecosystems,3–5 especially heavily calcified organisms, including corals.4,6 Indeed, coral reefs are one of the most sensitive ecosystems to this global stressor due to their delicate and complex physiology.7–11 The hydrocoral Millepora alcicornis shows a branching morphology and is one of the main corals to increase structural complexity on Brazilian reefs.12–14
Like scleractinian corals, hydrocorals feed heterotrophically on a variety of resources (mostly planktonic feeders). They also host unicellular dinoflagellate endosymbionts (i.e. zooxanthellae). Therefore, like scleractinian corals, they rely on a mutualistic symbiosis for autotrophic nutrition and calcification.15 In the scleractinian coral Mussismilia harttii, seawater acidification has been shown to negatively affect some key biochemical biomarkers involved in the coral calcification process and the photosynthetic metabolism of endosymbionts.16 Also, seawater acidification has been reported to induce oxidative stress and negatively affect the growth of the hydrocoral M. alcicornis.17,18 Therefore, as reported for scleractinian corals,9,10,16 hydrocorals are also prone to being affected by ocean acidification.5,17,19,20
In corals and hydrocorals, the calcification site pH is regulated relative to the surrounding seawater pH in order to promote the calcareous skeleton formation.8 However, the calcification site pH can be altered under conditions of ocean acidification, thus compromising several biochemical and physiological processes involved in the calcification of organisms.16,18,20 Results reported in the literature indicate that the magnitude of the seawater acidification effect on the growth of corals and hydrocorals is species-specific.4,21,22 In the hydrocoral M. alcicornis, a reduced growth rate was observed after long-term exposure to seawater acidification.18 At this point, it is important to note that a reduced seawater pH would lead to a similar reduced pH of the seawater present in the polyp's gastrovascular cavity.7 In this context, it is worth noting that pH regulation in the calcification medium is achieved by active proton (H+) pumping, an energy demanding process. Therefore, it is expected that this condition would lead to an increased energy requirement for maintenance of the coral calcification rate, thus resulting in a higher metabolic rate and reduced growth.21
In the present study, the effects of seawater acidification on the activities of hexokinase, pyruvate kinase, lactate dehydrogenase and citrate synthase were evaluated. It is worth noting that these enzymes catalyze irreversible key reactions involved in two major metabolic pathways, i.e., glycolysis and Krebs cycle. Therefore, the activities of these enzymes are finely regulated, as they are responsible for the control of these metabolic pathways in most organisms,23,24 including corals.25,26 Regarding glycolysis, the first and last steps of this metabolic pathway are regulated by hexokinase and pyruvate kinase, respectively. At the end of glycolysis, two molecules of ATP and two molecules of pyruvate are produced from one molecule of glucose.24
Under anaerobic conditions, pyruvate formed in the glycolytic pathway is converted into lactate, following a reaction catalyzed by lactate dehydrogenase.24 Under aerobic conditions, pyruvate formed in the glycolytic pathway is oxidized to acetyl-coenzyme A, which carries the carbon atoms of the acetyl group to the Krebs cycle for energy production. The first step of this metabolic pathway promotes the reaction of acetyl-coenzyme A and oxaloacetate, thus generating a molecule of citrate, with subsequent ATP, NADH and FADH2 production throughout the Krebs cycle. In the next step, the reduced coenzymes NADH and FADH2 are used by the electron transport chain to produce ATP through oxidative phosphorylation.24,27 It is important to note that the key reaction of acetyl-coenzyme A and oxaloacetate condensation mentioned above is catalyzed by citrate synthase.23,24
Considering the background described above, the aim of the present study was to evaluate the effects of reduced seawater pH on the activity of key enzymes involved in major energy metabolism pathways in the hydrocoral M. alcicornis. The enzymes evaluated included hexokinase, pyruvate kinase, lactate dehydrogenase and citrate synthase. It is expected that seawater acidification would lead to a reduced activity of enzymes involved in energy production, except for lactate dehydrogenase, since aerobic conditions are expected to occur over the whole period of the experiment by using the marine mesocosm system. In turn, a possible reduction in the activity of at least one of the metabolic enzymes under aerobic conditions would help to explain the previous reduced growth reported for the hydrocoral M. alcicornis under the same experimental conditions of seawater acidification tested in the present study.18
Materials and methods
Coral collection, acclimation and exposure to seawater acidification
Apical branches of four separate colonies of the hydrocoral M. alcicornis were collected in the Recife de Fora Municipal Marine Park (Porto Seguro, Bahia state, northeastern Brazil; S 16°24′37.3′′, W 38°59′02.2′′). Colonies sampled were at least 200 m apart. Hydrocoral branches collected (∼6 cm long) were fixed on ceramic plates with cyanoacrylate glue and acclimated for 20 days in the marine mesocosm system of the Coral Vivo Project (Arraial d'Ajuda, Porto Seguro). Therefore, branches collected at later experimental timepoints (0, 16 and 30 days) were already broken off at this initial stage. The branches from each colony were kept together in the same tank for the 20 day acclimation phase. However, branches from different colonies were kept in separate tanks in this initial phase. Hydrocorals fed only on natural foods present in the seawater.After the 20 day acclimation period, six branches (three branches for each exposure time: 16 and 30 days) from each original colony were randomly distributed in each of the four experimental tanks used for the four different pH treatments. Therefore, one separate tank was used for each colony subjected to each experimental condition tested (pH: 8.1 – control, 7.8, 7.5 and 7.2), thus totaling sixteen tanks. Therefore, four biological replicates were tested for each experimental condition, each one represented by branches from the same original colony exposed to the same experimental condition at each timepoint. In turn, technical replicates were represented by the three branches from each colony exposed to each experimental condition in the same tank at each timepoint.
The mesocosm structure and functioning were described in previous studies from our research group.16,18,20,28,29 Briefly, seawater, from the Mucugê reef located at 500 m from the research station, was used to mimic the abiotic characteristics and daily fluctuations of natural reefs in the marine mesocosm system. Reef seawater was continuously pumped into four 5000 L underground sumps. From there, seawater was again pumped to four 200 L elevated reservoirs, which were connected to the experimental tanks. The pH of the seawater from each of these reservoirs was continuously monitored and adjusted to reach the desired experimental conditions before reaching the experimental tanks through gravity. The experimental setup consisted of sixteen independent 100 L tanks. The seawater flux in each tank was adjusted to 8.3 L min−1 using a flow meter (TM 050; GPI, Wichita, KS, USA).
The methodology for seawater acidification in the present study was the same as previously reported by Marangoni et al. (2019).16 Briefly, mesocosm conditions were maintained at three reduced pH levels (7.8, 7.5 and 7.2) with respect to the ambient pH (8.1). Experimental pH conditions were selected considering predictions based on models of atmospheric CO2 emissions projected for years 2100 and 2300 under the IS92a scenario.2 Seawater acidification was achieved by bubbling CO2 into natural seawater. The seawater acidification process was controlled by means of a computerized system (Reef Angel® Controller, Dublin, CA, USA) coupled to pH electrodes (Gehaka 09RBCN, São Paulo, SP, Brazil). The pH sensors were calibrated every day, with pH 7.01 and pH 10.01 standard solutions (Hanna instruments, Woonsocket, RI, USA). In this case, the Reef Angel automatic calibration protocol was employed. Every week, the sensor's drift and response time were checked using a PG1800 Gehaka benchtop pH meter test protocol (Gehaka, São Paulo, SP, Brazil). In all cases, the free pH scale was employed.
The experiment was carried out under natural lighting. Light intensity (LI-COR, LI 250A Light Meter, LI-193 Underwater Spherical Quantum Sensor, Lincoln, NE, USA), pH (Gehaka 09RBCN pH electrodes, São Paulo, SP, Brazil), temperature and salinity (Instrutemp ITREF 10 optical refractometer, São Paulo, SP, Brazil) were monitored twice a day (08:00 a.m. and 08:00 p.m.). Every day (08:00 a.m.), seawater samples from each experimental tank were collected for measurements of total alkalinity and dissolved organic carbon concentration. Total alkalinity and dissolved organic carbon concentration were measured immediately after seawater sample collection. Total alkalinity was analyzed following Dickson et al. (2007)30 using an alkalinity titrator (AS-ALK2, Apollo SciTech Inc., Bogart, GA, USA) and certified reference material (ID: 132; Scripps Institute of Oceanography, La Jolla, CA, USA). In turn, dissolved organic carbon concentration was measured after seawater sample filtration (0.45 μm mesh filter) using a total organic carbon analyzer (TOC analyzer 5050A, Shimadzu, Japan). Levels of pCO2 and aragonite saturation state (Ωar) values were calculated for each treatment. Calculations were performed based on the total alkalinity and mean pH data using CO2calc. version 1.2.9 software. The pCO2 values were 351.8 μatm, 939.0 μatm, 1683.4 μatm, and 3494.3 μatm for pH 8.1, pH 7.8, pH 7.5 and pH 7.2, respectively. In turn, the Ωar values corresponded to 3.89, 1.96, 1.23, and 0.64, respectively.
After the 20 day period of hydrocoral acclimation to the mesocosm condition, twelve branches of M. alcicornis (three branches of each of the four hydrocoral colonies) were collected, individually placed into a cryovial and immediately frozen in liquid nitrogen. Therefore, three technical replicates (three branches of each colony) were collected per tank, while four biological replicates (branches of the four different colonies) were collected and frozen as described above. Samples collected were further maintained in an ultra-freezer (−80 °C) until laboratory analysis. The remaining branches were exposed to the control condition or to the three different seawater acidification treatments, for 16 and 30 days. Three branches of each of the four colonies were tested per tank, totaling twelve hydrocoral branches for each replicate at each timepoint. Four independent replicates (four raceway tanks) were tested for each experimental condition (control or acidification treatment). During the experiment, the hydrocorals fed only on natural foods present in the seawater. A qualitative color gradient ranging from six (darkest tone) to zero (white color), adapted from a Coral Health Chart (https://coralwatch.org/), was used to monitor variation in hydrocoral tissue color. Under all experimental conditions, no clear sign of bleaching was observed over the whole experimental time. After the exposure period, hydrocoral samples were collected, frozen and stored as described above for acclimated hydrocorals.
In the laboratory, samples (hydrocoral branches) were macerated in liquid nitrogen, using a mortar and pestle. Macerated samples were divided into aliquots, conditioned in cryovials and immediately frozen in liquid nitrogen. Frozen samples were stored in an ultra-freezer (−80 °C) until enzyme activity analysis.
Enzyme assays
The activities of glycolytic and Krebs cycle enzymes were analyzed according to Lallier and Walsh (1991),31 and Lauer et al. (2012),32 with slight modifications. Each sample aliquot (∼0.5 g) was homogenized in ice-cold buffer (50 mM imidazole; 0.1 mM PMSF; pH 7.8), using a sonicator (Qsonica Model CL-188, Newtown, CT, USA). The homogenate was then centrifuged (10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 × g; 20 min; 4 °C) and the supernatant was used as the enzyme source.
000 × g; 20 min; 4 °C) and the supernatant was used as the enzyme source.
Enzyme activities were measured spectrophotometrically using a microplate reader (ELx808IU, BioTek Instruments, Winooski, VT, USA). Glycolytic enzyme (hexokinase, pyruvate kinase and lactate dehydrogenase) assays were performed using an imidazole buffer solution (75 mM; pH 7.4). Enzyme activity was measured based on NAD+/NADH oxidation/reduction by following the sample absorbance at 340 nm. Krebs cycle enzyme (citrate synthase) assay was performed using a HEPES buffer solution (50 mM; pH 8.1). Enzyme activity was measured based on 5-5′-dithio-2-nitrobenzoic (DTNB) reduction by following the sample absorbance at 412 nm. Specific conditions were used for each enzyme assay, as follows (final concentrations): hexokinase (150 mM MgCl2, 10 mM D-glucose, 1.2 mM NAD+, 120 U mL−1 glucose-6-phosphate dehydrogenase from Leuconostoc mesenteroides and 60 mM ATP); pyruvate kinase (150 mM MgCl2, 750 mM KCl, 37.5 mM ADP, 3 mM NADH, 1500 U mL−1 lactate dehydrogenase and 5 mM phosphoenolpyruvate); lactate dehydrogenase (7.7 mM pyruvate and 3 mM NADH); and citrate synthase (2.3 mM acetyl coenzyme A, 3.5 mM DTNB and 3.5 mM oxaloacetate). All assays were performed at 25 °C. For each experimental treatment, three branches of each hydrocoral colony were analyzed, thus totaling twelve branches per pH condition (pH 8.1, pH 7.8, pH 7.5, and pH 7.2) at each experimental time (0, 16 and 30 days). Protein content in the sample homogenate was determined based on the absorbance at 550 nm, using the Bradford reagent (Sigma-Aldrich, St. Louis, MO, USA). Data were expressed as enzyme units (EU) per mg protein (UE per mg protein).
Data presentation and statistical analysis
Data on seawater physicochemical parameters (pH, temperature and salinity: n = 60; alkalinity and dissolved organic carbon: n = 30) and enzyme activity (n = 4, i.e., four coral colonies; three branches of each coral colony were measured at each exposure time) were expressed as mean ± standard deviation. Data on seawater physicochemical parameters were compared by one-way analysis of variance (ANOVA). Enzyme activity data after hydrocoral acclimation to the mesocosm conditions and after exposure to the control pH condition (ambient seawater pH: 8.1) for 16 and 30 days were compared by one-way ANOVA. In turn, enzyme activity data after exposure to different experimental conditions for 16 and 30 days were subjected to two-way (exposure time and pH treatment) ANOVA. Post-hoc significant differences among the different experimental treatments were evaluated using Fisher's Least Significant Difference (LSD) test. ANOVA assumptions, i.e., data normality and homogeneity of variances, were previously verified using the normal probability plot for raw residuals and the Cochran C test, respectively. In all cases, the significance level adopted was 95% (α = 0.05). All statistical analyses were performed using Statistica 7.0 software (StatSoft, Tulsa, OK, USA).Results
Data on seawater chemistry are presented in Table 1. The results obtained showed that pH treatments were successfully achieved and maintained throughout the experimental period. No significant difference was observed among replicates for the same experimental condition (p > 0.05; one-way ANOVA). However, a significant difference was observed among the different experimental conditions tested (p < 0.05; one-way ANOVA, followed by the LSD test). General mean values (± standard deviation) were 8.10 ± 0.096 (nominally pH 8.1; pH range: 7.71–8.34), 7.74 ± 0.185 (nominally pH 7.8; pH range: 6.60–8.41), 7.51 ± 0.160 (nominally pH 7.5; pH range: 6.22–8.36), and 7.21 ± 0.190 (nominally pH 7.2; pH range: 6.15–8.35). No significant difference in light intensity, temperature, salinity, alkalinity and dissolved organic carbon concentration was observed among the different replicates for the same experimental condition, as well as among the different experimental conditions (p > 0.05; one-way ANOVA). General mean values for the whole experimental period corresponded to 285.95 ± 39.13 μmol photons per m2 per s, 25.12 ± 0.30 °C, 35.04 ± 0.38, 117.00 ± 1.77 mg CaCO3 per L, and 3.35 ± 0.35 mg C per L, respectively.| Parameter | Experimental condition | ||||
|---|---|---|---|---|---|
| pH 8.1 | pH 7.8 | pH 7.5 | pH 7.2 | ||
| pH | 8.10 ± 0.096A | 7.74 ± 0.185B | 7.51 ± 0.160C | 7.20 ± 0.190D | |
| Temperature | 25.19 ± 0.03A | 25.00 ± 0.03A | 25.16 ± 0.03A | 25.13 ± 0.04A | |
| Salinity | 35.11 ± 0.39A | 35.02 ± 0.41A | 35.13 ± 0.37A | 35.07 ± 0.34A | |
| Alkalinity | 119.2 ± 1.7A | 117.1 ± 1.5A | 115.8 ± 2.6A | 115.9 ± 1.5A | |
| DOC | 3.43 ± 0.37A | 3.56 ± 0.36A | 3.20 ± 0.31A | 3.19 ± 0.34A | |
| Light intensity | 285.0 ± 41.3A | 296.4 ± 41.5A | 285.5 ± 33.9A | 277.0 ± 39.8A | |
Data on enzyme activities in hydrocoral collected after the 20 day period of acclimation are presented in Table S1 (ESI).† For each enzyme, no significant difference was observed among mean values for the four hydrocoral colonies (p > 0.05; one-way ANOVA). Data on enzyme activities in hydrocoral collected after the 16 day and 30 day periods of exposure are presented in Tables S2 and S3, respectively (ESI).† For each enzyme, no significant difference was observed (p > 0.05; one-way ANOVA) among the mean values for the four hydrocoral colonies for each pH condition (pH 8.1, 7.8, 7.5, and 7.3) at each exposure time (16 and 30 days). Therefore, mean values for each pH condition and exposure time were calculated based on the twelve branches analyzed (three branches of each of the four hydrocoral colonies).
In hydrocorals kept at ambient seawater pH (8.1), no significant change (p > 0.05; one-way ANOVA) was observed in enzyme activity [hexokinase (Fig. 1), pyruvate kinase (Fig. 2), lactate dehydrogenase (Fig. 3) and citrate synthase (Fig. 4)] over the experimental period (0, 16 and 30 days). In hydrocorals exposed to seawater acidification for 16 days, no significant change (p > 0.05; two-way ANOVA) was observed in hexokinase (Fig. 1), pyruvate kinase (Fig. 2), lactate dehydrogenase (Fig. 3) and citrate synthase (Fig. 4) activity. In hydrocorals exposed to seawater acidification for 30 days, no significant change (p > 0.05; two-way ANOVA) was observed in hexokinase (Fig. 1), pyruvate kinase (Fig. 2) and lactate dehydrogenase (Fig. 3) activity. However, citrate synthase activity was markedly inhibited (p < 0.05; two-way ANOVA, followed by the LSD test) in hydrocorals exposed to seawater acidification treatments (Fig. 4).
 | ||
| Fig. 1 Hexokinase activity (UE per mg protein) in the hydrocoral Millepora alcicornis kept under the control condition (ambient seawater: pH 8.1) or exposed to different levels of CO2-driven seawater acidification (pH 7.8, 7.5 and 7.2) for 16 and 30 days in a marine mesocosm system. Data are expressed as mean ± standard deviation (n = 12). Different capital letters indicate significantly different mean values (p < 0.05; one-way ANOVA, followed by the LSD test) under the control condition (pH 8.1) at the different experimental times (0, 16 and 30 days). Different lowercase letters indicate significantly different mean values (p < 0.05; two-way ANOVA, followed by the LSD test) among the experimental conditions (exposure time and pH treatment). | ||
 | ||
| Fig. 2 Pyruvate kinase activity (UE per mg protein) in the hydrocoral Millepora alcicornis kept under the control condition (ambient seawater: pH 8.1) or exposed to different levels of CO2-driven seawater acidification (pH 7.8, 7.5 and 7.2) for 16 and 30 days in a marine mesocosm system. Data are expressed as mean ± standard deviation (n = 12). Different capital letters indicate significantly different mean values (p < 0.05; one-way ANOVA, followed by the LSD test) under the control condition (pH 8.1) at the different experimental times (0, 16 and 30 days). Different lowercase letters indicate significantly different mean values (p < 0.05; two-way ANOVA, followed by the LSD test) among the experimental conditions (exposure time and pH treatment). | ||
 | ||
| Fig. 3 Lactate dehydrogenase activity (UE per mg protein) in the hydrocoral Millepora alcicornis kept under the control condition (ambient seawater: pH 8.1) or exposed to different levels of CO2-driven seawater acidification (pH 7.8, 7.5 and 7.2) for 16 and 30 days in a marine mesocosm system. Data are expressed as mean ± standard deviation (n = 12). Different capital letters indicate significantly different mean values (p < 0.05; one-way ANOVA, followed by the LSD test) under the control condition (pH 8.1) at the different experimental times (0, 16 and 30 days). Different lowercase letters indicate significantly different mean values (p < 0.05; two-way ANOVA, followed by the LSD test) among the experimental conditions (exposure time and pH treatment). | ||
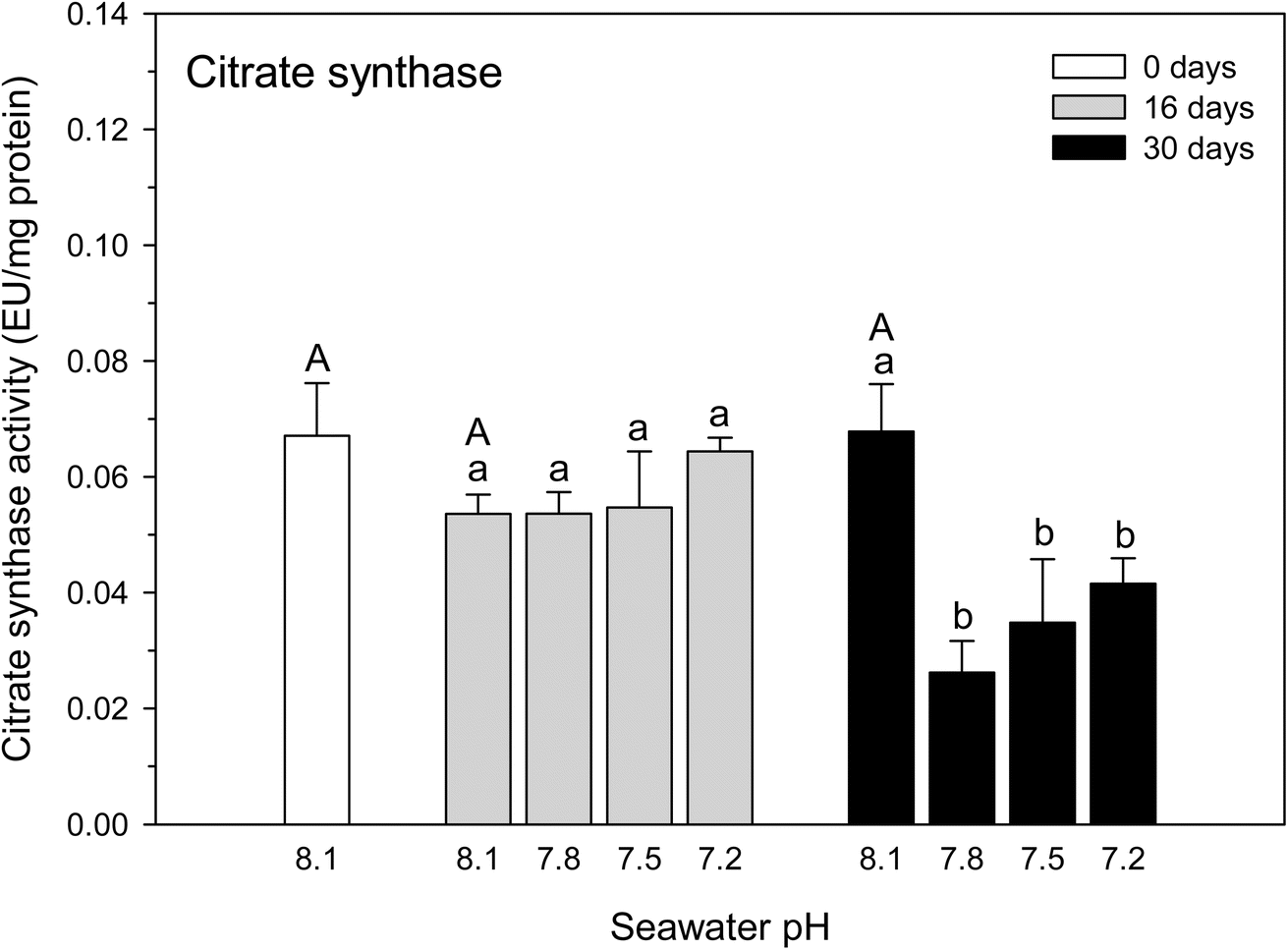 | ||
| Fig. 4 Citrate synthase activity (UE per mg protein) in the hydrocoral Millepora alcicornis kept under the control condition (ambient seawater: pH 8.1) or exposed to different levels of CO2-driven seawater acidification (pH 7.8, 7.5 and 7.2) for 16 and 30 days in a marine mesocosm system. Data are expressed as mean ± standard deviation (n = 12). Different capital letters indicate significantly different mean values (p < 0.05; one-way ANOVA, followed by the LSD test) under the control condition (pH 8.1) at the different experimental times (0, 16 and 30 days). Different lowercase letters indicate significantly different mean values (p < 0.05; two-way ANOVA, followed by the LSD test) among the experimental conditions (exposure time and pH treatment). | ||
Discussion
Based on the hypothesis outlined for the present study, the reduced growth rate previously observed in the hydrocoral M. alcicornis could be associated with lower energy production related to the reduced activities of key enzymes linked to glycolysis and the Krebs cycle. Therefore, we would expect the activities of the tested enzymes to decrease under an ocean acidification scenario. However, our data reported show that CO2-driven seawater acidification, as low as pH 7.1, did not affect hexokinase and pyruvate kinase activities, even after 30 days of exposure to this environmental stressor. This finding indicates that the pyruvate formation rate through the glycolytic pathway was likely not affected after M. alcicornis exposure to the different levels of seawater acidification tested.Regarding the lack of response of the lactate dehydrogenase, it is important to note that this enzyme plays a crucial role, especially under anaerobic conditions. In fact, pyruvate formed in the glycolytic pathway is converted into lactate, following a reaction catalyzed by this enzyme.24 Therefore, the lack of the seawater acidification effect on lactate dehydrogenase activity observed in the present study is evidence that hydrocorals maintained aerobic metabolism throughout the whole experimental period.
Differently from the other enzymes analyzed, the activity of the citrate synthase was markedly reduced in the hydrocoral M. alcicornis exposed to any of the seawater acidification conditions (pH 7.8, 7.5 and 7.2) for 30 days. Therefore, aerobic energy production was certainly compromised in M. alcicornis chronically exposed to seawater acidification. The reduced citrate synthase activity together with the lack of change in lactate dehydrogenase activity observed in hydrocoral exposed to seawater acidification in the 30 day exposure indicates that M. alcicornis was likely not able to sustain an energy production steady state. This finding can thus explain, at least in part, the reduced growth previously reported for this hydrocoral species.18 This statement is in line with other biochemical and molecular studies on corals subjected to seawater acidification. In fact, Edmunds and Wall (2014)33 reported that protein synthesis was negatively affected by high pCO2 in recruits of the scleractinian coral Seriatopora caliendrum, with consequences on aerobic respiration. In turn, Kaniewska et al. (2012)34 reported that genes involved in the Krebs cycle were downregulated in the scleractinian coral Acropora millepora exposed to pH 7.8–7.9 and 7.6–7.7 for 28 days. Also, Vidal-Dupiol et al. (2013)21 reported that exposure to pH ≤ 7.4 for 21 days induced a downregulation of genes coding for proteins involved in coral calcification, as well as heterotrophic and autotrophic related proteins in the scleractinian coral Pocillopora damicornis. More recently, Glazier et al. (2020)10 reported a lowered expression of genes involved in the energy metabolism of the scleractinian coral Lophelia pertusa exposed to pH 7.6 for six months in comparison to those kept at ambient pH (7.9).
From an ecological perspective, our findings suggest the possibility of ocean acidification impacts on reef-builder corals. Indeed, it was recently demonstrated that two natural coral reef communities on the back reef of Mo'orea (French Polynesia) showed a decreased net calcification of the natural coral reef community subjected to a high pCO2 condition (949 μatm) compared to that exposed to the ambient pCO2 condition (393 μatm).35 In the context of global changes, it is interesting to note that citrate synthase activity was also shown to be inhibited in the hydrocoral M. alcicornis and the scleractinian corals Mussismilia hispida and Porites branneri subjected when exposed to combined seawater acidification and increased temperature (pH 7.8 and 28.5 °C) compared to the control conditions (pH 8.1 and 26.0 °C).36 Therefore, the inhibition of citrate synthase activity induced by reduced seawater pH, as demonstrated in the present study with the hydrocoral M. alcicornis and scleractinian corals,36 can be a good biomarker of the reduced growth observed in hydrocorals and scleractinian corals after chronic exposure to ocean acidification.16,18 Indeed, it was demonstrated that the hydrocoral M. alcicornis is capable of coping with acidic conditions only for a short period of time (16 days). However, it shows clear negative responses (oxidative stress) after chronic exposure (30 days) to seawater acidification, which could compromise hydrocoral health.17 Interestingly, we also found a significant and marked reduction in the citrate synthase activity only after 30 days of M. alcicornis exposure to seawater acidification. Based on these findings, it is expected that both corals and hydrocorals would negatively respond to ocean acidification over the long term, showing compromised health, a lower growth rate and gross production.
Conclusion
The observed lack of changes in the activity of hexokinase and pyruvate kinase provides some evidence that the glycolytic pathway in the hydrocoral M. alcicornis is functioning normally even after 30 days of exposure to seawater acidification (pH 7.8, 7.5 and 7.2). In turn, the observed inhibition of the citrate synthase activity indicates that the first step of the Krebs cycle for energy production is compromised, leading to a reduced activity of citrate synthase in the hydrocoral M. alcicornis. This would be implicated in a reduced energy availability to sustain an adequate metabolic rate. Such a condition could thus explain, at least in part, the reduced growth previously reported in this hydrocoral species subjected to seawater acidification for 30 days. Therefore, future work would be focused on the establishment of a direct connection between lower energy production through the Krebs cycle and the reduced growth in hydrocorals and corals long-term exposed to an ocean acidification scenario.Data availability
The data that support the findings of this study are available on request from the corresponding author, [AB].Author contributions
Conceptualization: Andrea Carlina Jesulich, Laura Fernandes de Barros Marangoni, Joseane Aparecida Marques, Cristiano Macedo Pereira, Gustavo Adolpho Santos Duarte, Débora de Oliveira Pires, Clovis Barreira e Castro, Emiliano Nicolas Calderon, and Adalto Bianchini. Data curation: Andrea Carlina Jesulich, Mariana Machado Lauer, Laura Fernandes de Barros Marangoni, Joseane Aparecida Marques, and Yuri Dornelles Zebral Adalto Bianchini. Formal analysis: Andrea Carlina Jesulich, Mariana Machado Lauer, Laura Fernandes de Barros Marangoni, Joseane Aparecida Marques, Yuri Dornelles Zebral, and Adalto Bianchini. Funding acquisition: Débora de Oliveira Pires, Clovis Barreira e Castro, and Adalto Bianchini. Investigation: Andrea Carlina Jesulich, Mariana Machado Lauer, Laura Fernandes de Barros Marangoni, Joseane Aparecida Marques, Cristiano Macedo Pereira, Gustavo Adolpho Santos Duarte, Débora de Oliveira Pires, Clovis Barreira e Castro, Emiliano Nicolas Calderon, and Adalto Bianchini. Methodology: Andrea Carlina Jesulich, Mariana Machado Lauer, Laura Fernandes de Barros Marangoni, Joseane Aparecida Marques, Yuri Dornelles Zebral, Cristiano Macedo Pereira, Gustavo Adolpho Santos Duarte, Débora de Oliveira Pires, Clovis Barreira e Castro, Emiliano Nicolas Calderon, and Adalto Bianchini. Project administration: Débora de Oliveira Pires, Clovis Barreira e Castro, Emiliano Nicolas Calderon, and Adalto Bianchini. Resources: Andrea Carlina Jesulich, Mariana Machado Lauer, Laura Fernandes de Barros Marangoni, Joseane Aparecida Marques, Yuri Dornelles Zebral, Cristiano Macedo Pereira, Gustavo Adolpho Santos Duarte, Débora de Oliveira Pires, Clovis Barreira e Castro, Emiliano Nicolas Calderon, and Adalto Bianchini. Supervision: Débora de Oliveira Pires, Clovis Barreira e Castro, Emiliano, and Adalto Bianchini. Writing – original draft: Andrea Carlina Jesulich, Mariana Machado Lauer, Laura Fernandes de Barros Marangoni, Joseane Aparecida Marques, Yuri Dornelles Zebral, Cristiano Macedo Pereira, Gustavo Adolpho Santos Duarte, Débora de Oliveira Pires, Clovis Barreira e Castro, Emiliano Nicolas Calderon, and Adalto Bianchini. Writing – review & editing: Yuri Dornelles Zebral and Adalto Bianchini.Conflicts of interest
There are no conflicts of interest to declare.Acknowledgements
This study was financially supported by the International Development Research Centre (IDRC, Ottawa, Canada; grant # 104519-003), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES – Programa Ciências do Mar, Brasília, DF, Brazil; grant # 84/2010) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq – Instituto Nacional de Ciência e Tecnologia de Toxicologia Aquática, Brasília, DF, Brazil; grant # 573949/2008-5). We would like to thank the Rede de Pesquisas Coral Vivo, Petrobras and Arraial d’Ajuda Eco Parque for all logistical assistance provided during the mesocosm experiment. A. Bianchini (Proc. # 311410/2021-9) is a research fellow from the Brazilian CNPq and was supported by the International Canada Research Chair Program (IDRC). M.M. Lauer was a postdoctoral fellow from the Brazilian CNPq. A.C. Jesulich was supported by the Scholarship Organization of American States (OAS) and PAEC-OEA-GCUB 2013 programs.References
- J.-P. Gattuso, A short summary of the current knowledge on ocean acidification, in. Bridging the Gap between Ocean Acidification Impacts and Economic Valuation: Regional Impacts of Ocean Acidification on Fisheries and Aquaculture, ed. N. Hilmi, D. Allemand, C. Kavanagh, D. Laffoley, M. Metian, D. Osborn and S. Reynaud, International Union for Conservation of Nature and Natural Resources, Gland, 2015, pp. 19–21 Search PubMed.
- Core Writing Team, H. Lee and J. Romero, Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, International Panel on Climate Change, Geneva, 2023 Search PubMed.
- C. A. Logan, A review of ocean acidification and America's response, BioScience, 2010, 60, 819–828 CrossRef.
- K. J. Kroeker, L. R. Kordas, R. Crim, I. E. Hendriks, R. Ramajo, G. S. Singh, C. M. Duarte and J.-P. Gattuso, Impacts of ocean acidification on marine organisms: quantifying sensitivities and interaction with warming, Global Change Biol., 2013, 19, 1884–1896 CrossRef PubMed.
- J. M. Lough and M. J. H. van Oppen, Introduction: Coral Bleaching – Patterns, Processes, Causes and Consequences, in Coral Bleaching – Patterns, Processes, Causes and Consequences, ed. M. J. H. van Oppen and Lough J. M., Springer, Cham, 2nd edn, 2018, 1, pp. 1–8 Search PubMed.
- Y. D. Zebral, J. S Fonseca, J. A. Marques and A. Bianchini, Carbonic anhydrase as a biomarker of global and local impacts: insights from calcifying animals, Int. J. Mol. Sci., 2019, 20, 3092 CrossRef CAS PubMed.
- M. Holcomb, A. A. Venn, E. Tambutté, S. Tambutté, D. Allemand, J. Trotter and M. McCulloch, Coral calcifying fluid pH dictates response to ocean acidification, Sci. Rep., 2014, 4, 1–4 Search PubMed.
- A. A. Venn, The role of pH regulation in coral symbiosis and biomineralization, Presented in Part at 4th Coral Reef Ecology Symposium, Bremen, June, 2014 Search PubMed.
- R. Albright, Ocean acidification and coral bleaching, in Coral Bleaching – Patterns, Processes, Causes and Consequences, ed. M. J. H. van Oppen and J. M. Lough, Springer, Cham, 2nd edn, 2018, 12, pp. 295–323 Search PubMed.
- A. Glazier, S. Herrera, A. Weinnig, M. Kurman, C. E. Gómez and E. Cordes, Regulation of ion transport and energy metabolism enables certain coral genotypes to maintain calcification under experimental ocean acidification, Mol. Ecol., 2020, 29, 1657–1673 CrossRef CAS PubMed.
- K. H. Tisthammer, E. Timmins-Schiffman, F. O. Seneca, B. L. Nunn and R. H. Richmond, Physiological and molecular responses of lobe coral indicate nearshore adaptations to anthropogenic stressors, Sci. Rep., 2021, 11, 3423 CrossRef CAS PubMed.
- E. O. C. Coni, C. M. Ferreira, R. L. Moura, P. M. Meirelles, L. Kaufman and R. B. Francini-Filho, An evaluation of the use of branching fire-corals (Millepora spp.) as refuge by reef fish in the Abrolhos Bank, eastern Brazil, Environ. Biol. Fishes, 2013, 96, 45–55 CrossRef.
- I. C. S. Leal, P. H. C. Pereira and M. E. de Araújo, Coral reef fish association and behaviour on the fire coral Millepora spp. in north-east Brazil, J. Mar. Biol. Assoc. U. K., 2013, 93, 1703–1711 CrossRef.
- A. L. Luza, J. P. Quimbayo, C. E. L. Ferreira, S. R. Floeter, R. B. Francini-Filho, M. G. Bender and G. O. Longo, Low functional vulnerability of fish assemblages to coral loss in Southwestern Atlantic marginal reefs, Sci. Rep., 2022, 12, 1–12 CrossRef PubMed.
- C. E. Dubé, C. A. F. Bourmaud, A. Mercière, S. Planes and E. Boissin, Ecology, biology and genetics of Millepora hydrocorals on coral reefs, in. Invertebrates - Ecophysiology and Management, ed. S. Ray, G. Diarte-Plata and R. Escamilla-Montes, IntechOpen, London, 2020, vol. 2, pp. 1–36 Search PubMed.
- L. F. B. Marangoni, M. M. A. N. Pinto, J. A. Marques and A. Bianchini, Copper exposure and seawater acidification interaction: Antagonistic effects on biomarkers in the zooxanthellate scleractinian coral Mussismilia harttii, Aquat. Toxicol., 2019, 206, 123–133 CrossRef CAS PubMed.
- D. C. Luz, Y. D. Zebral, R. D. Klein and A. Bianchini, Oxidative stress in the hydrocoral Millepora alcicornis exposed to CO2-driven seawater acidification, Coral Reefs, 2018, 37, 571–579 CrossRef.
- L. F. B. Marangoni, E. M. Calderon, J. A. Marques, G. A. S. Duarte, C. M. Pereira, C. B. Castro and A. Bianchini, Effects of CO2-driven acidification of sea water on the calcification process in the calcareous hydrozoan Millepora alcicornis (Linnaeus, 1758), Coral Reefs, 2017, 36, 1133–1141 CrossRef.
- J. B. Lewis, The biology and ecology of the hydrocoral Millepora on coral reefs, Adv. Mar. Biol., 2006, 50, 1–55 Search PubMed.
- L. F. B. Marangoni, J. A. Marques and A. Bianchini, Fisiologia de corais: a simbiose coral-zooxantela, o fenômeno de branqueamento e o processo de calcificação, in. Conhecendo os Recifes Brasileiros – Rede de Pesquisas Coral Vivo, ed. C. Zilberberg, A. Pinto, J. A. Marques, L. Feitosa and L. Marangoni, Museu Nacional, Rio de Janeiro, 2016, vol. 4, pp. 55–72 Search PubMed.
- J. Vidal-Dupiol, D. Zoccola, E. Tambutté, C. Grunau and C. Cosseau, Genes related to ion-transport and energy production are upregulated in response to CO2-driven pH decrease in corals: new insights from transcriptome analysis, PLoS One, 2013, 8, 1–11 CrossRef PubMed.
- D. Brown and P. J. Edmunds, Differences in the responses of three scleractinians and the hydrocoral Millepora platyphylla to ocean acidification, Mar. Biol., 2016, 163, 1–10 CrossRef CAS.
- H. Morrison, Enzyme Activity Sites and Their Reaction Mechanisms, Academic Press, London, 2021 Search PubMed.
- D. L. Nelson and M. M. Cox, Lehninger: Principles of Biochemistry, W. H. Freeman, New York, 2021 Search PubMed.
- A. Sánchez-Quinto and L. I. Falcón, Metagenome of Acropora palmata coral rubble: Potential metabolic pathways and diversity in the reef ecosystem, PLoS One, 2019, 14, e0220117 CrossRef PubMed.
- L. B. Linsmayer, D. D. Deheyn, L. Tomanek and M. Tresguerres, Dynamic regulation of coral energy metabolism throughout the diel cycle, Sci. Rep., 2020, 10, 19881 CrossRef CAS PubMed.
- D. F. Wilson, Oxidative phosphorylation: regulation and role in cellular and tissue metabolism, J. Physiol., 2017, 595, 7023–7038 CrossRef CAS PubMed.
- G. Duarte, E. N. Calderon, C. M. Pereira, L. F. B. Marangoni, H. F. Santos, R. S. Peixoto, A. Bianchini and C. B. Castro, A novel marine mesocosm facility to study global warming, water quality, and ocean acidification, Ecol. Evol., 2015, 5, 4555–4566 CrossRef PubMed.
- V. C. Sarmento, T. P. Souza, A. M. Esteves and P. J. P. Santos, Effects of seawater acidification on a coral reef meiofauna community, Coral Reefs, 2015, 34, 955–966 CrossRef.
- A. G. Dickson, C. L. Sabine and J. R. Christian, Guide To Best Practices For Ocean CO2 Measurements, North Pacific Marine Science Organization, Sidney, 2007 Search PubMed.
- F. H. Lallier and P. J. Walsh, Metabolic potential in tissues of the blue crab, Callinectes sapidus, Bull. Mar. Sci., 1991, 48, 665–669 Search PubMed.
- M. M. Lauer, C. B. Oliveira, N. L. I. Yano and A. Bianchini, Copper effects on key metabolic enzymes and mitochondrial membrane potential in gills of the estuarine crab Neohelice granulata at different salinities, Comp. Biochem. Physiol., C, 2012, 156, 140–147 CAS.
- P. J. Edmunds and C. B. Wall, Evidence that high pCO2 affects protein metabolism in tropical reef corals, Biol. Bull., 2014, 227, 68–77 CrossRef CAS PubMed.
- P. Kaniewska, P. R. Campbell, D. I. Kline, M. Rodriguez-Lanetty and D. J. Miller, Major cellular and physiological impacts of ocean acidification on a reef building coral, PLoS One, 2012, 7, 1–12 CrossRef PubMed.
- S. S. Doo, P. J. Edmunds and R. C. Carpenter, Ocean acidification effects on in situ coral reef metabolism, Sci. Rep., 2019, 9, 12067 CrossRef PubMed.
- M. S. Angonese, S. C. Faria and A. Bianchini, Is citrate synthase an energetic biomarker for Southwestern Atlantic corals? A comparative, biochemical approach under simulated climate changes, Coral Reefs, 2022, 41, 213–222 CrossRef.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d4va00083h |
| This journal is © The Royal Society of Chemistry 2025 |