 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceConvergence in biomineralization patterns across animal eggshells†
Gerben Debruyn *a,
Seung Choib,
Jessica L. Dobsona,
Yana Maudensc,
Karen De Clerckc,
Matthew D. Shawkeya,
Shukang Zhangd and
Liliana D’Alba*e
*a,
Seung Choib,
Jessica L. Dobsona,
Yana Maudensc,
Karen De Clerckc,
Matthew D. Shawkeya,
Shukang Zhangd and
Liliana D’Alba*e
aDepartment of Biology, Evolution and Optics of Nanostructures Group, Ghent University, Karel Lodewijk Ledeganckstraat 35, 9000 Gent, Belgium. E-mail: gerben.debruyn@ugent.be
bResearch Institute of Basic Sciences, Seoul National University, Seoul 08826, South Korea
cCenter for Textile Science and Engineering, Faculty of Engineering and Architecture, Ghent University, Tech Lane Science Park 70a, B-9052 Ghent, Belgium
dKey Laboratory of Vertebrate Evolution and Human Origins, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China
eNaturalis Biodiversity Center, Darwinweg 2, 2333 CR Leiden, Netherlands. E-mail: liliana.dalbaaltamirano@naturalis.nl
First published on 26th February 2025
Abstract
Shelled eggs are key components of animal reproduction on land, evolving independently in distant lineages of terrestrial animals including nematodes, gastropods, annelids, arthropods and chordates. They perform critical functions such as the exchange of gases between embryo and the environment, desiccation avoidance and protection from harmful radiation, microbial infection and mechanical damage. A core mechanism behind eggshell multifunctionality is the incorporation of biominerals (mainly calcium carbonate and calcium phosphate) into the shell. Very little is known about eggshell structure in invertebrates, but some recent pioneering studies have proposed that similar mineralization patterns may have evolved convergently in eggshells of pulmonate gastropods, some insects, and vertebrates. However, because a detailed characterization of the structural and chemical composition of invertebrate eggshells is not available, it has not been possible to test this hypothesis. Here, we use computed tomography, electron microscopy, electron backscatter diffraction analyses, atomic force microscopy, spectroscopy, and histochemistry to characterize and compare microstructure and chemical composition of pulmonate gastropod, insect and vertebrate eggshells. These techniques revealed the universal presence of an organic matrix in mineralized eggshells. However, disparities in the distribution of calcium throughout the shell, crystallographic orientation that appears random in invertebrates (but not vertebrates), and presence of different calcium types including the rare and unstable vaterite highlight divergence whose functional significance should be the subject of future study.
Introduction
Eggs are remarkable structures that nurture and protect animal embryos, serving as self-contained units that provide everything needed for embryonic development. The earliest eggs appeared more than 500 million years ago,1 and over time, eggs diversified in size, shape and composition.2–5 One of the most remarkable advancements in the evolution of eggs is the development of a protective outer covering, the eggshell, which is almost exclusive to terrestrial species.3,6 Eggshells enabled animals to exploit a vast array of habitats by rendering the egg impermeable while mediating critical functions including the exchange of gases between the embryo and the environment, desiccation avoidance and protection from harmful radiation, microbial attack and mechanical damage.7–10A key mechanism behind these functions is the incorporation of calcareous minerals (e.g. carbonate, phosphate and oxalate) into the eggshell.11–15 Calcification occurs exclusively in terrestrial eggs and is assumed to provide the ultimate protective barrier against desiccation and damage,15,16 although empirical evidence to support this hypothesis has only recently begun to emerge.8,17,18
The form and composition of eggshells vary widely across animals, yet many evolutionary lineages independently evolved calcified eggs.2,11,13,19,20 Eggshell calcification took place convergently in pulmonate gastropods, arthropods and amniote vertebrates as early as 300 million years ago with the invasion of land2,3,21,22 (Fig. 1). Several lineages also independently reduced their thick calcified shell (e.g. marine turtles).6,19 Previous research has shown that, when present, eggshell mineralization may occur within a spectrum ranging from the simple accumulation of calcium ions to the deposition of crystals or discrete layers within the shell, to an almost complete calcification of eggshells (as in birds and crocodiles).11,14,21,23–25
 | ||
| Fig. 1 Occurrence and general plan of mineralized eggshells across animal taxa. (A) Simplified, phylum level schematic tree of bilaterian animals showing the production of shelled (cleidoic) eggs (E) in animals parallels terrestrial lifestyles (T). At least 3 groups of land animals evolved calcified (Ca) eggshells: vertebrates, insects and mollusks. In annelids, strong accumulation of calcium ions has been detected in a few species.26 (B) Photographs of taxon-representative eggs and egg cases used in this study (not shown to scale); top to bottom: birds (Chordata: Psittacula roseata), reptiles (Chordata: Pelusios carinatus), stick insects (Insecta: Mearnsiana bullosa), cockroaches (Insecta: Periplaneta americana), snails (Gastropoda: Stylommatophora). (C) Schematic drawings showing generalized morphology of eggshells. Cuticle (Cu), calcified layer (CL), mammillary knob (M), fibers (F), porous layer (PL), dark layer (D), laminar layer (Lm), middle layer (Md), calcium crystal (Cy), inner membrane (IM), outer layer (OL). | ||
Models of eggshell biomineralization
The ultrastructure and chemical composition of eggshell mineralization varies extensively across taxa, yet significant gaps exist in our understanding of these processes. While the ultrastructure of vertebrate eggshells, particularly in birds, has been studied extensively, research on mineralized invertebrate eggshells has lagged, despite their importance in both ecological and applied fields.Biomineralization is a complex but ubiquitous process by which minerals are deposited in tissues of living organisms. Particularly for avian eggshells, two competing models have been proposed to explain mineralization. A first mechanism proposes that calcified columnar units often observed in eggshells arise from the competition in crystal growth between neighbouring nucleation sites.27,28 An alternative mechanism proposes biological regulation and the involvement of organic components in structuring crystal formation.29–31
This biogenic model of mineralization states that the presence of an organic matrix (composed of proteins, polysaccharide, and proteoglycans) orchestrates the structural and crystallographic organization of the mineral phase. It is well known that numerous mineralized structures in invertebrates, often made of calcium carbonate—such as skeletons and storage deposits—develop from the transformation of initially disordered precursors, such as amorphous calcium carbonate (ACC).32–34 Precursors are delivered to confined calcification sites, where they aggregate and form crystals through particle attachment crystallization. This process is facilitated by an organic interface that acts as a scaffold for mineralization. Specifically in birds, the eggshells consistently exhibit a purely calcitic composition with a polycrystalline configuration.35 It is now accepted that mineralization of the avian eggshell is under biological control, where the organic matrix components modulate crystal growth and determine the size and orientation of calcite crystals.35,36 The process begins with the deposition of substantial amounts of ACC particles onto the mammillary knobs (organic aggregates and the sites of nucleation of calcite crystal). This ACC accumulation transitions into aggregates of calcite microcrystals oriented in all directions, which gradually expand both vertically through competitive growth and through the elongation of crystals parallel to the surface, modulated by matrix proteins. The initially adjacent crystalline units become fused and calcification is arrested when a thin organic layer (cuticle) is deposited. Proteins of the matrix form a fibrillar network throughout the eggshell, from the mammillary knobs to the calcitic units (where the matrix fibers become perpendicularly arranged)29 and into the vertical crystal layer (where the fibers are also vertically aligned).34
Considerable progress has been made in elucidation of the process of eggshell mineralization using avian eggs as models.16,37–40 More recently, a renewed interest in explaining the vast diversity of animal eggshell morphologies and the relationships between the organic and the mineral phases more broadly has stimulated studies on non-avian species.41–44 However, our understanding of the process of eggshell biomineralization, particularly in invertebrates, is still limited. This paper aims to examine key structural and compositional features of various, distantly related animal eggshells and to explore similarities and disparities in their biomineralization. We test the hypothesis that animal groups share a convergent mechanism of eggshell biomineralization based on an organic matrix-controlled model.
Results and discussion
Structure and distribution of minerals within eggshells among different animal taxa
Complementary microscopy techniques including electron backscatter diffraction (EBSD), traditional histology, and electron microscopy shows that calcification of almost all the examined arthropod, mollusk, and vertebrate eggshells occur in well-defined layers. The exception to this pattern is the cockroach ootheca. However, the structure and distribution of calcified deposits varies notably within and between eggshells of different animal taxa (Fig. 1). The eggshell of bird and non-avian reptile (hereafter referred to as reptiles) is highly ordered, with calcium carbonate often forming a thick layer on top of a fibrous proteinaceous membrane (Fig. 2A, B, D and E). Calcium is organized into a hierarchical structure initially formed by the binding of amorphous calcium carbonate (ACC) to the mammillary knobs,35 which act as nucleation sites and anchoring points between the mineralized part of the eggshell and the organic membrane. The palisade layer lies over the mammillary layer and comprises the bulk of the eggshell (two-thirds) in the form of large, columnar fused calcium crystals. This layer sometimes transitions into a thinner vertical crystal layer, as in palaeognaths such as rheas.45 The cuticle, the outermost layer in contact with the environment, is usually present in avian eggshells, but was not discernible in the eggshells we analyzed. | ||
| Fig. 2 False colored SEM images of animal eggs. (A) Psittacula roseata (Aves), (B) Rhea americana (Aves), (C) Pomacea canaliculata (Gastropoda), (D) Caiman crocodilus (Reptilia), (E) Pelusios carinatus (Reptilia), (F) Achatina fulica (Gastropoda), (G) Mearnsiana bullosa (Phasmatodea), (H) Heteropteryx dilatata (Phasmatodea), (I) Periplaneta lateralis (Blattodea). Red false colouring indicates the location of the mineralized portions of the shell as informed by the complementary techniques (EBSD, EDX and differential staining) used in this study. | ||
By contrast, calcified eggshells are less common in invertebrates, found here only in arthropods and gastropods. These exhibit considerable variation: the mineralization patterns of the four snail species analyzed here appear entirely different and surpass the diversity seen in avian eggshells.13 For example, the eggs of Helix pomatia consisted of discrete individual, unfused crystalline units (ESI Fig. 1†). The eggshells of Achatina fulica and Archachatina marginata were both composed of fused granules. By contrast, Pomacea canaliculata eggs are laid in clusters in which individual eggs are cemented together by a mass of non-fused calcified vaterite spherules roughly 3 μm in size (ESI Fig. 2†). In different species from the same genus (P. paludosa),46 vaterite spherules are also present both in the surface of each egg and in the cement between eggs.46
Eggs or ootheca of arthropods, including stick insects, cockroaches and mantids, are all calcified to some degree.11,12,47 However, the distributions of minerals within these structures are fundamentally different. In the cockroach Periplaneta lateralis distinctive crystals spread across the oothecal wall. These crystals appear randomly distributed and range from 5 to 30 μm (Fig. 2I). While the exact function of these crystals is not known, it is hypothesized that they increase the hardiness of the eggs and/or facilitate the tanning of the ootheca by creating a more acidic environment.11 Stick insect eggshells, on the other hand, have a clearly stratified structure (including the exo- and endochorion). The layer closest to the embryo (endochorion) is not mineralized and therefore excluded from our samples. However, certain parts of the exochorion are mineralized. The exochorion consists of different layers depending on the species. Generally, the phasmid eggshell can be divided into three layers, but variable depending on the author.12,48–50 The outermost layer (consisting of the porous layer and dark layer; Fig. 1) and the inner membrane (near the endochorion) are not mineralized. The layers in between are strongly calcified. In M. bullosa we observed clear calcium granules in these calcified layers (Fig. 2G). For most phasmid species such as Rhynchacris ornata, and Heteropteryx dilatata these units are less obvious and rather transition from a continuous layer to one forming pillars with flamelike structures (Fig. 2H).
Invertebrate eggshells exhibit more variation in crystallographic orientation, morphology and arrangement
Our crystallographic analyses identified the presence of at least two layers in avian and crocodile eggshells (Fig. 3A and B) suggesting that calcite deposition occurs in at least two different steps.51 Turtle eggshells show a single layer, i.e. the mammillae knobs continuously extend to the main calcified layer. Complex superficial sculptures or lacunae are present in some crocodile species such as Caiman (Fig. 3C). These lacunae likely form as a result of dissolution processes that progress from the surface toward the mammilla.19 Most dinosaur eggs and non-avian tetanuran theropod eggshells are composed of single layers, but those of maniraptoran dinosaur (a clade including oviraptorosaurs and modern birds) and crocodilian eggs are characterized by at least two layers.45,51,52 The crystallographic arrangement is similar for archosaurs: there is an initial mineral phase near the mammillary knobs, containing more randomly oriented grains, followed by the growth of columnar calcareous units with preferred vertical orientation throughout the outer portion of the eggshell.53,54 This alignment was strongest in the eggshell of Rhea americana (Fig. 3B). It is still weakly present in the other species such as the parakeet Psittacula roseata, (Fig. 3A) and, also observed in the caiman (Fig. 3C). In the turtle eggshell, aragonite crystals originate from the organic core and expand radially without forming a columnar layer (Fig. 3D). Specific orientations of the crystals can improve the mechanical properties of the eggshells (e.g. Guinea fowl eggshells)55 by increasing compressive strength, which would reduce the risk of fracture when the incubating parent sits on the eggs or when eggs are buried in compact soils.18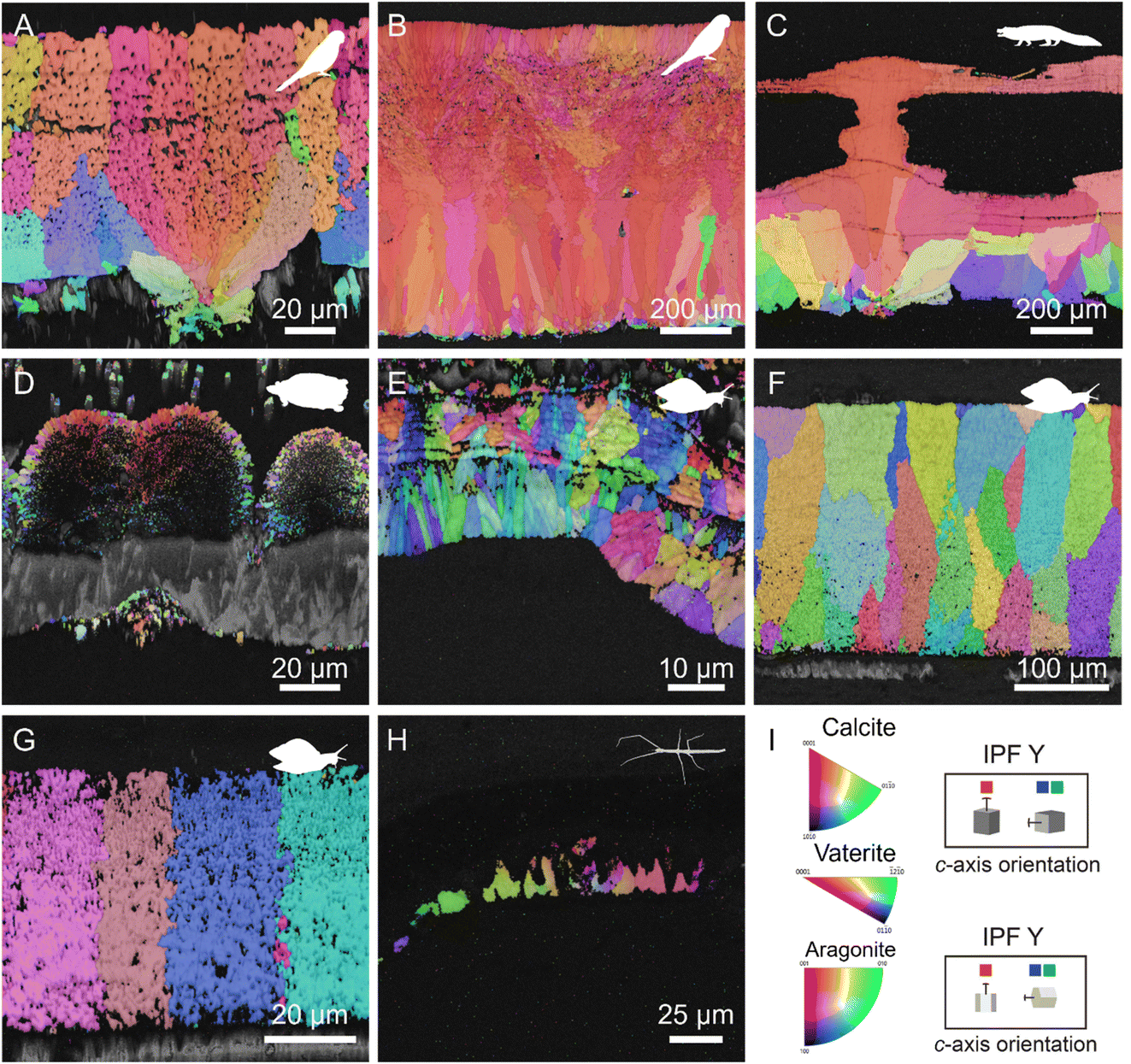 | ||
| Fig. 3 EBSD maps of radially sectioned eggshells. Inverse pole figure (IPF) maps displaying the crystalline structure, and orientation of the calcium carbonate crystals in the shells. (A) Psittacula roseata (Aves), (B) Rhea americana (Aves), (C) Caiman crocodilus (Reptilia). Except for the turtle (D) Pelusios carinatus (Reptilia) that presents the aragonite polymorph, all other vertebrates show calcite crystals. In invertebrates, (E) shows a rare occurrence of vaterite in the eggshell of Pomacea canaliculata (Gastropoda) while the other snails (F) Archachatina marginata (Gastropoda) and (G) Achatina fulica (Gastropoda) have calcite. (H) Clear calcite crystals were also visible in the stick insect Heteropteryx dilatata (Insecta). (I) Legends for inverse pole figure Y maps and a schematic representation of c-axis orientation for calcium carbonate polymorphisms: calcite and vaterite for which the crystals are hexagonal and aragonite with an orthorhombic shape. | ||
Notably, the crystallographic orientation and arrangement for invertebrate eggshells is completely different (Fig. 3E–H). Generally, no layering is present within the calcified layer of the snail eggshells (Fig. 3F and G), which are composed of a single stack. In the eggshell of the Pomacea canaliculata, two potential layers can be distinguished, the inner with long, narrow crystal grains that are preferentially oriented and the outer one with randomly oriented crystals (Fig. 3E). This observation aligns with Tompa’s (1976) description,13 which noted the presence of two or three stacked layers of calcitic crystals varying in size and orientation (i.e. Anguispira alternata, another species of pulmonate land snail). The inner surface of the snail eggshells differs from mammillary knobs of vertebrate eggshells in that their calcium carbonate grains are relatively large compared to the thickness of eggshell and differ consistently throughout the eggshell as in vertebrate eggshells. Perhaps the lack of well-organized nucleation sites (as in vertebrate eggshells) results in a more random distribution and growth of crystals. For example, mammillary knobs (homoplastic with that of birds) were observed in the eggshells of the giant South American snail, Megalobulimus oblongus; interestingly, only in that species, were regularly distributed columnar calcitic units also observed superjacent to the knobs.13 Alternatively, the organic matrix, which in bird and caiman eggshells is more regular and organized than invertebrates (discussed below), could facilitate a more ordered crystal growth, as in the highly ordered nacre in bivalves, where a structured network of organic rings predetermines the arrangement and final size of individual aragonite crystal units or tablets.56 Calcite grains are present in the middle of the stick insect eggshells (Fig. 3H), but do not extend to the inner or outer end of eggshells and do not show a preferred orientation. The shape and distribution of calcitic crystals are dissimilar in the two species of stick insects, suggesting that calcite distribution varies highly between stick insects. Polarized microscopy revealed two mineralized layers, and our FTIR data suggest that one of those layers is calcium oxalate while the EBSD (Fig. 3H) shows that the other is calcite.12,57 However, we did not detect the presence of calcium oxalate using EBSD.
Most carbonate indexed by EBSD analysis in this study is calcite. However, we detected aragonite in turtle eggshells (Fig. 3D),23,58,59 and vaterite in the eggshell of the snail P. canaliculata (Fig. 3E). As in stick insect eggs, we found no calcium oxalate (due to the absence of Kikuchi bands when the grain was analyzed) in the cockroach ootheca using EBSD. However, extraction of crystals has demonstrated calcium oxalate crystals in these egg cases.11 The crystals appear to be at a lower (topographic) height relative to the surrounding matrix (ESI Fig. 3†), possibly preventing the crystals from exposure during polishing, and obscuring Kikuchi bands detection. Thus, although EBSD found no signals from the cockroach and stick insect eggshells, other techniques strongly suggest the presence of calcium oxalate in these eggshells.
Interplay between organic matrix and mineral phase is convergent across animal eggshells
We reveal, across a range of animal groups, that biomineralization in eggshells consistently involves the co-occurrence of an organic matrix and minerals (Fig. 4). In all taxa, the mineral component (i.e. different types of calcium compounds) is found in close association with an organic phase (i.e. organic matrix) and seems critical to the formation and structuring of mineralized eggshells. This may represent a convergent evolutionary mechanism of eggshell calcification. It also argues against the hypothesis that mineralization is determined solely through crystal growth competition (i.e. inorganic control). Rather, we propose that the formation of organic nucleation sites and scaffolds plays a pivotal role in the process. The presence of organic matrices is well documented in bird eggshells and in other biomineralized structures in invertebrates such as the calcified shell of sea urchins and mollusks, where they regulate mineral deposition, influence crystal growth and orientation and delimit the connection between individual crystals and/or crystal layers.60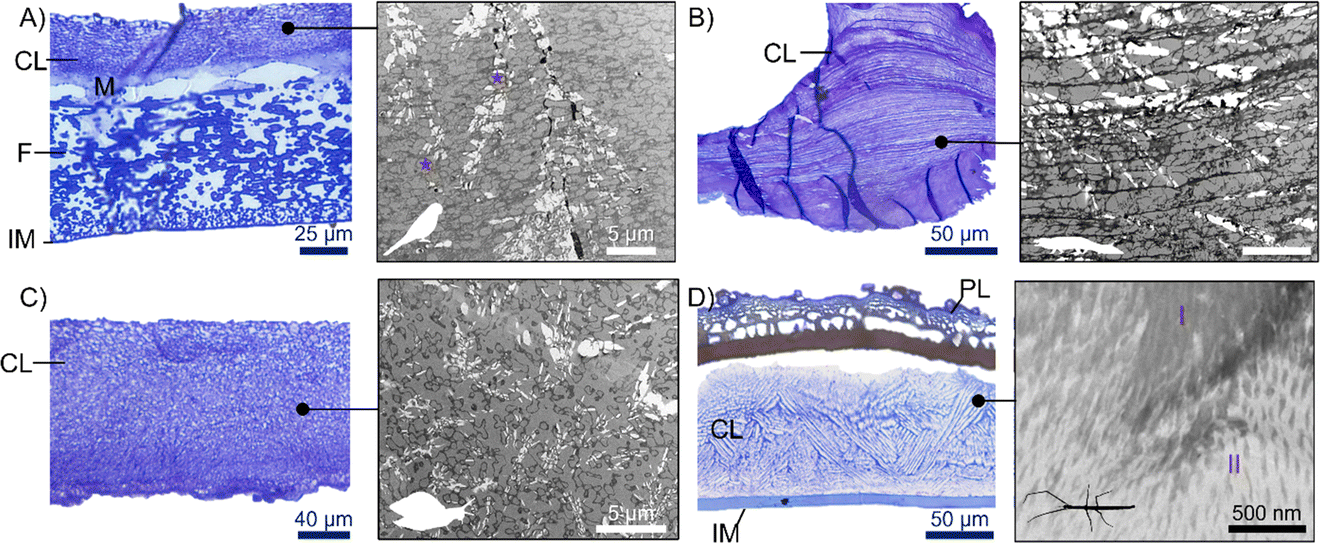 | ||
| Fig. 4 Micrographs of the organic matrix of animal eggshells. Blue panels: toluidine blue staining (highlighting the general structure of eggshells) and, grey panels: stained TEM micrographs (revealing organic matrix morphology). (A) Psittacula roseata (Aves) with imprint of vesicles (a few indicated by stars), (B) Caiman crocodilus (Reptilia), (C) Achatina fulica (Gastropoda), (D) Heteropteryx dilatata (Insecta) with two visible different organic matrix regions identified by the higher and lower electron density in the TEM image (I and II respectively). Calcified layer (CL), mammillary knob (M), fibers (F), inner membrane (IM), porous layer (PL). Please note, long white voids in the TEM images are artifacts. | ||
Various hypotheses have been proposed for the role of proteins and polysaccharides in biogenic crystal formation. For example, the organic matrix can be a substrate that binds calcium and therefore works strictly as a nucleation site for the growth of inorganic crystals. This is seen in the formation of nacres in pearl oyster Pinctada fucata.61 In the ootheca of Periplaneta lateralis we observed a thin, fibrillar organic mesh at the periphery of each of the polygonal crystals and potentially also interspersed across the polygonal crystals (Fig. 5 and ESI Fig. 3†). In the turtle eggshell (Pelusios carinatus) we also observed a network of radially arranged organic fibrils underneath and across the mammillary cores that may serve as the nucleation site of the aragonite crystals (Fig. 3D)62 but we failed to visualize the extension of organic fibrils throughout the calcareous units towards the outermost shell surface (ESI Fig. 4†).
 | ||
| Fig. 5 Details of Periplaneta lateralis eggshell microstructure. (A) Toluidine blue stained histology cross section. (B) TEM image of a cross section. (C) EDX micrograph shows in bright blue the presence of calcium, overlaid with SEM image of the cross section of the eggshell. (D) TEM of a calcium pocket with fibrillar network at the periphery with inset of the fibrillar network. | ||
In all other species, we found that the organic matrix is present throughout the entire mineralized area, and not only near the nucleation site, or between calcium crystals. Every shell contains a distinct fused mineralized layer, organic network in the form of a lattice or mesh of rectangular or polygonal spaces acting as scaffolding basal to the crystalline phase (Fig. 4).
In birds, we found that the organization of the organic matrix is variable along the eggshell, beginning with a dense packing closer to the nucleation sites at the mammillary knobs, then becoming progressively less compact throughout the palisades (Fig. 4A). In the chicken eggshell, the organic fibrils become vertically aligned coincidentally with the vertical layer of the calcified shell.63 In stick insects the organic matrix is morphologically divided in two layers that vary in fibril density and coincide with the transition between two calcareous layers previously described12 (Fig. 4D).
Interestingly, calcium carbonate is present in the eggshell of the snail Pomacea canaliculata both as a distinctive layer where it co-occurs with an organic matrix and as scattered spherulites that are embedded in an organic matrix similar to the mineralization process proposed for the shells of mussels Pinna nobilis and Mytilus edulis.64 While the organic matrix is always present, its overall structure differs between taxa. The scaffolding tends to be a reticulate mesh in vertebrates; in the caiman shell, fine horizontal fibers mirror the accretion lines seen in the calcified units (Fig. 4B) and in the eggshell of Psittacula roseata the matrix also showed the imprint of vesicles (Fig. 4A). This organic matrix seemed to be organized differently in the invertebrates. In snails, the organic matrix has a homogeneous, bicontinuous pattern (Fig. 4C). In stick insects, there seems to be some organization in the organic matrix that changes in orientation throughout the calcified layer (Fig. 4D).
Presence of acid proteoglycans and proteins within organic matrix guide calcium deposition
A large number of chemicals with control of specific aspects of biomineralization have been described recently: negatively charged and containing carboxylate, sulfate, and phosphate, which bind Ca2+ ions.65,66 One particular group, proteoglycans (i.e. acid mucopolysaccharides) consisting of a protein core with negatively charged linear polysaccharide side chains, consistently recur in biomineralization of calcium carbonate in bird eggs. In birds and reptiles, mineralization initiates with nucleation of calcium on mammillae that possess proteoglycans with sulfated mucopolysaccharides.37,40,67 Although we did not observe nucleation sites for crystal growth in snails, cockroaches and stick insects, we did find that mucopolysaccharides, specifically acid mucopolysaccharides, were omnipresent along with the calcified layers in all species (Fig. 6). This strongly suggests an essential role in biomineralization of animal eggs, binding the calcium from the initial to the end of the mineralizing process. Moreover, as is evident from the stick insect eggshells, acid mucopolysaccharides might be important in the biomineralization of both calcium carbonate and calcium oxalate, where they are also pervasively present (Fig. 6D).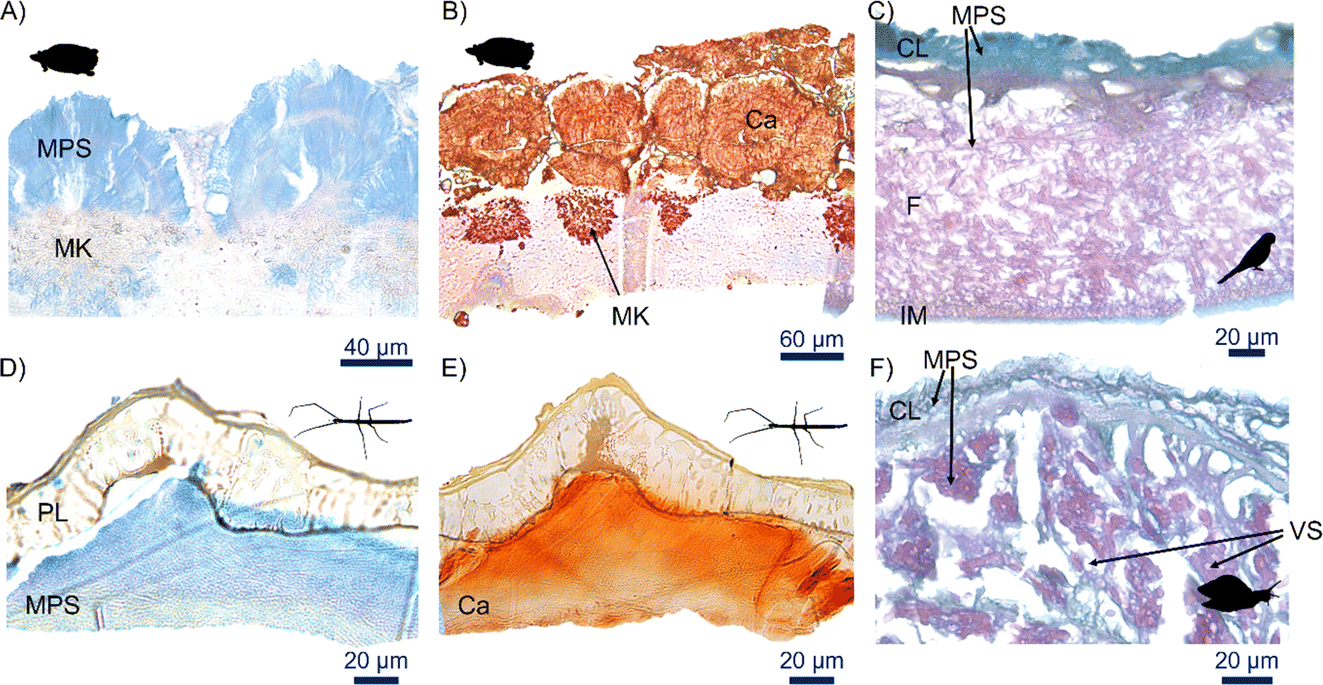 | ||
| Fig. 6 Occurrence of mucopolysaccharide in calcified layers. Stained histology cross section of eggshells for Pelusios carinatus (turtle) (A) showing acid mucopolysaccharide (MPS) stained by Alcian blue in blue and (B) calcium stained by Alizarin red in red. Combined Alcian blue/PAS staining for (C) Psittacula roseata (bird) showing acid mucopolysaccharide in blue and neutral mucopolysaccharides in magenta. Note that the calcified layer is incomplete due to decalcification protocol. (D) Cross section of Rhynchacrys ornata (stick insect) stained with Alcian blue showing acid mucopolysaccharides in blue and (E) alizarin red that stains red for calcium. (F) Pomacea canaliculata (snail) cross sections showing acid mucopolysaccharide in blue and neutral mucopolysaccharides in magenta. Mammillary knobs (MK), porous layer (PL), membrane fibers (F), inner membrane (IM), vaterite spherulites (VS). | ||
Disparities in mineral composition between animal eggshells
Although distantly related species likely evolved similar mechanisms of eggshell mineralization – most likely involving the facilitated accumulation and guided control of calcium deposition and crystal organization by acid mucopolysaccharides in the proteinaceous matrix – the specific types of constituent minerals vary widely within and between taxa. While birds predominantly incorporate calcium carbonate in the form of calcite (a highly stable polymorph) as shown by the typical calcite peaks of 1393, 874 and 713 cm−1 other animals incorporate different forms of calcium carbonate (i.e. vaterite and aragonite, Fig. 3),23 and even different types of calcium compounds (Fig. 7). Most reptiles also use calcite, except for turtles that integrate aragonite instead; typical aragonite peaks are visible around 1455, 854, 712, and 700 cm−1 (Fig. 7B). Aragonite is less stable (metastable), but it could be beneficial as it is denser than calcite, and aragonite eggshells are on average stiffer than calcite ones.54 Invertebrates also incorporate various forms of calcium minerals in their eggshells, e.g. stick insects use calcium carbonate (calcite) and calcium oxalate. FTIR showed that stick insect eggs only have calcium oxalate with the typical peaks around 1612, 1309, and 776 cm−1.57 However, in agreement with previous reports for the group,12 our EBSD results indicated that phasmid eggshells also possess calcium carbonate in the form of calcite (Fig. 3H). | ||
| Fig. 7 Fourier transform infra-red spectra of the calcified eggs: (A) birds (red hues) with typical calcite peaks, (B) reptiles (orange hues) with different calcium carbonate polymorphs: calcite for Caiman crocodilus (crocodile) and aragonite for Pelusios carinatus (turtle). (C) Arthropods (green hues): stick insects (H. dilatata and M. bullosa) and cockroaches (Periplaneta lateralis) both showing typical calcium oxalate peaks. (D) Snails in blue hues show calcium carbonate in calcite polymorphism for A. fulica and vaterite polymorphism for Pomacea canaliculata. | ||
Calcium oxalate is also present in the cockroach ootheca. While the oxalate peaks were visible with FTIR analysis, these could have been masked by other peaks, such as those of chitin (also evident with FTIR) and perhaps due to the relatively high amount of organic material we did not find a strong chemical fingerprint of calcium oxalate which is known to be present in the ootheca of many cockroaches.11 Interestingly, both cockroaches and stick insects possess calcium oxalate This could be a conserved mechanism to harden tissues among arthropods11,12 or could be linked to their life history. Stick insects, and to a lesser extent cockroaches, feed mainly on plants which usually produce high concentrations of calcium oxalate as an anti-herbivory mechanism.68 The high availability (i.e. supersaturation) of oxalate salts relative to calcium carbonate precursors may preferentially bind to calcium, mirroring the process by which kidney stones form when oxalate-rich diets are eaten in combination with inadequate hydration.69 This can also explain why phasmids possess both calcium carbonate (calcite) and calcium oxalate within their eggshell. A similar transition in the incorporation of calcium minerals is seen in reptiles, which sometimes have calcite instead of aragonite depending on whether they are sampled in the wild or kept captive.70
Calcium composition in snails was variable, we found calcite in Achatina fulica, and vaterite (an uncommon and metastable polymorph of calcium carbonate) in Pomacea canaliculata. Preferentially incorporating vaterite could be advantageous to snails as vaterite forms easily in different environments (e.g. in high pH),71 and may require less energy to make when mineralization occurs rapidly.
Conclusion
Throughout animal evolution, egg size increased as species adapted to life on land. Larger eggs hold more water, but also require stronger shells, so mineralized eggshells likely evolved multiple times in gastropods, arthropods, and vertebrates as they colonized terrestrial environments. Whether these evolutionary events involved similar or distinct mechanisms has been an open question largely due to a lack of morphological data on invertebrate eggshells.Here we show that biomineralization among these diverse eggs seems to occur through similar underlying mechanisms, but with different chemistry. All species studied use an organic matrix as a scaffolding or anchoring structure to guide initial deposition of calcium and influence the growth of crystals. Vertebrates and invertebrates might use different types of polysaccharides and proteins that have similar functional properties. This strong convergence suggests that the use of an organic matrix is a highly advantageous mechanism of biomineralization, and future research should focus on understanding its advantages, and on how the variation in egg chemistry between species affects egg functionality.
Experimental
Sample collection and preparation
We collected eggs from different private breeders (ESI Table 1†). We first cleaned the eggshells with distilled water using a sonicator to remove egg yolk residues, then dried them at room temperature and cut them into sections for the subsequent analysis.Further, we embedded untreated and decalcified eggshell pieces in Epon resin using the procedure as described in Dunning et al.72 and for the paraffin embedding decalcified samples are rinsed twice in phosphate buffered saline (PBS) for 30 min each. Next, a dehydration step was performed through a series of ethanol washes (30%, 50%, 70%) of 1 hour each, followed by three 1 hour washes in 100% ethanol and two 1 hour washes in xylene. The samples are then put in paraffin at 58 °C for three 1 hour sessions and then put into paraffin blocks. After this the sample blocks get trimmed and cut, to the necessary thickness that each technique requires, using a Leica UC-6 microtome (Leica Microsystems, Germany).
Scanning- and transmission electron microscopy (SEM, TEM)
To obtain SEM images we mounted eggshell sections to an aluminum stub with the cut surface facing up, allowing a clear cross-sectional view, and sputter-coated with gold–palladium (10–12 nm thick). SEM micrographs were taken using a FlexSEM 1000 (Hitachi, Japan).For TEM we followed previously published protocols,73 where untreated Epon blocks are cut into ultra-thin sections (100 nm) and transferred to formvar-coated copper grids, stained with 1% uranyl acetate and lead citrate and viewed on a JEM-1010 (JEOL Ltd, Japan).
The sample blocks get trimmed and the decalcified Epon blocks were cut to 100 nm thick for TEM, and both the Epon (decalcified and untreated) and paraffin (decalcified) blocks were cut to 3 μm thick sections for traditional histology using a Leica UC-6 ultramicrotome (Leica Microsystems, Germany).
Histological staining and light microscopy
For traditional light microscopy all blocks were cut to 3 μm thick sections. We used the untreated Epon sections for the alizarin red S staining, which stains for calcite, amorphous calcium carbonate, hydroxyapatite, aragonite, and other calcium phosphates and oxalates. We used the decalcified Epon embedded eggshell sections for toluidine blue staining to visualize the overall eggshell structure. The decalcified paraffin sections were used for the combined Alcian blue and Periodic Acid-Schiff (PAS) staining, which stains for mucopolysaccharides.Fourier transform infrared spectroscopy
We used Fourier transform infrared (FTIR) spectroscopy to obtain the chemical fingerprint of the eggshells. The eggshell sections are pulverized using a pestle and mortar after which the FTIR analysis was performed using a Nicolet iS50 FTIR spectrometer combined with a universal Attenuated Total Reflectance attachment (Thermo Fisher Scientific, USA). For each sample, we collected and took the average of 32 scans (4 cm−1 resolution) within the 4000–400 cm−1 range.Electron backscatter diffraction (EBSD)
We used electron backscatter diffraction (Symmetry; Oxford Instruments) to obtain microstructural and crystallographic information of eggshells with good biomineral crystallinity. We mainly followed established EBSD protocols for eggshell analysis and data presentation.74 Briefly, the eggshell sections embedded in resin were cut and then lapped with sandpaper with decreasing grit sizes. The lapped surfaces were further polished with diamond paste and colloidal silica to make perfectly polished sections. The prepared EBSD specimens were tilted to 70° in a SEM, and then analyzed under 25.0 mm working distance, acceleration voltage of 15 kV, and emission current of 92.8 μA. The data were represented in inverse pole figure Y maps that effectively show the c-axis directions of minerals.Data availability
The authors confirm that the data supporting the findings of this study are available within the article and its supplementary information.†Author contributions
Conceptualization: GD, LD, SC, SZ, KDC; methodology: GD, SC, LD, KDC, SZ; investigation: GD, SC, JD, YM, SZ, LD; visualization: GD, SC, JD, YM, SZ, LD; supervision: LD, KDC, MDS; writing – original draft: GD, SC, LD; writing – review & editing: all authors.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This work was supported by FWO grant GOA7921N, AFOSR grant FA9550-23-1-0622, and EOARD grant FA8655-23-2-7041. We want to sincerely thank Phillipe Jouk, Curator Aquarium, Reptielen en zeezoogdieren at the Antwerp Zoo, and Catherine Vancsok, Scientific Direction of the Pairi Daiza Foundation for their support by providing access to the reptile eggshells used in the study. We also want to thank Mieke Soenens (Ghent University) for making the histological sections, Tine Herman for the use of her stick insect egg photographs, and the Transmission Electron Microscopy Core Facility at UGent for access to their equipment, without which the study would not have been possible. We thank Yuxia Zhu and Xun Jin (Institute of Vertebrate Paleontology and Paleoanthropology) for assistance in the making of thin sections and help with SEM imaging, respectively, and Yong Park (Seoul National University) for his advice for EBSD map representation.References
- S. C. Morris, Eggs and embryos from the Cambrian, BioEssays, 1998, 20(8), 676–682 CrossRef CAS PubMed.
- G. C. Packard and M. J. Packard, Evolution of the cleidoic egg among reptilian antecedents of birds, Am. Zool., 1980, 20(2), 351–362 CrossRef.
- M. Mazzini, G. Callaini and C. Mencarelli, A comparative analysis of the evolution of the egg envelopes and the origin of the yolk, Ital. J. Zool., 1984, 51(1–2), 35–101 Search PubMed.
- H. E. Hinton, Insect eggshells, Sci. Am., 1970, 223(2), 84–91 CrossRef CAS PubMed.
- T. A. Rawlings, Adaptations to physical stresses in the intertidal zone: the egg capsules of neogastropod molluscs, Am. Zool., 1999, 39(2), 230–243 CrossRef.
- L. J. Legendre, D. Rubilar-Rogers and G. M. Musser, et al., A giant soft-shelled egg from the Late Cretaceous of Antarctica, Nature, 2020, 583(7816), 411–414 CrossRef CAS PubMed.
- D. W. Zeh, J. A. Zeh and R. L. Smith, Ovipositors, amnions and eggshell architecture in the diversification of terrestrial arthropods, Q. Rev. Biol., 1989, 64(2), 147–168 CrossRef.
- Y. Chang and P. Y. Chen, Hierarchical structure and mechanical properties of snake (Naja atra) and turtle (Ocadia sinensis) eggshells, Acta Biomater., 2016, 31, 33–49 CrossRef PubMed.
- M. J. Packard, G. C. Packard and T. J. Boardman, Structure of Eggshells and Water Relations of Reptilian Eggs, Herpetologica, 1982, 38(1), 136–155 Search PubMed . http://www.jstor.org/stable/3892368.
- L. D’Alba and M. D. Shawkey, Mechanisms of antimicrobial defense in avian eggs, J. Ornithol., 2015, 156, 399–408, DOI:10.1007/s10336-015-1226-1.
- B. Stay, A. King and L. M. Roth, Calcium oxalate in the oothecae of cockroaches, Ann. Entomol. Soc. Am., 1960, 53(1), 79–86 CrossRef CAS.
- A. Moscona, Studies of the egg of Bacillus libanicus (Orthoptera, Phasmidae) I. The egg envelopes, J. Cell Sci., 1950, 3(14), 183–193 CrossRef.
- A. S. Tompa, A comparative study of the ultrastructure and mineralogy of calcified land snail eggs (Pulmonata: Stylommatophora), J. Morphol., 1976, 150(4), 861–887 CrossRef PubMed.
- L. D’Alba, J. Goldenberg and A. Nallapaneni, et al., Evolution of eggshell structure in relation to nesting ecology in non-avian reptiles, J. Morphol., 2021, 282(7), 1066–1079 CrossRef PubMed.
- M. T. Hincke, Y. Nys, J. Gautron, K. Mann, A. B. Rodriguez-Navarro and M. D. McKee, The eggshell: structure, composition and mineralization, Front. Biosci.-Landmark, 2012, 17(4), 1266–1280 CrossRef CAS PubMed.
- Y. Dauphin, G. Luquet, A. Perez-Huerta and M. Salomé, Biomineralization in modern avian calcified eggshells: similarity versus diversity, Connect. Tissue Res., 2018, 59(sup1), 67–73 CrossRef CAS PubMed.
- K. Tanaka, D. K. Zelenitsky and F. Therrien, Eggshell porosity provides insight on evolution of nesting in dinosaurs, PLoS One, 2015, 10(11), e0142829 CrossRef PubMed.
- A. B. Rodríguez-Navarro, N. Domínguez-Gasca and D. Athanasiadou, et al., Guinea fowl eggshell structural analysis at different scales reveals how organic matrix induces microstructural shifts that enhance its mechanical properties, Acta Biomater., 2024, 178, 244–256 CrossRef PubMed.
- H. H. Schleich and W. Kästle, Reptile egg-shells, SEM atlas, published online, 1988 Search PubMed.
- A. Moscona, Utilization of Mineral Constituents of the Egg-shell by the Developing Embryo of the Stick Insect, Nature, 1948, 162(4106), 62–63 CrossRef CAS PubMed.
- R. R. Kohring, Reflections on the origin of the amniote egg in the light of reproductive strategies and shell structure, Hist. Biol., 1995, 10(3), 259–275 CrossRef.
- M. R. Lee, M. E. Hodson and G. N. Langworthy, Crystallization of calcite from amorphous calcium carbonate: earthworms show the way, Mineral. Mag., 2008, 72(1), 257–261 CrossRef CAS.
- G. Debruyn, J. Geltmeyer and E. Schoolaert, et al., Hydric environment and chemical composition shape non-avian reptile eggshell absorption, Integr. Comp. Biol., 2024, 64, 107–119 CrossRef CAS PubMed.
- D. C. Deeming, Nesting environment may drive variation in eggshell structure and egg characteristics in the Testudinata, J. Exp. Zool., Part A, 2018, 329(6–7), 331–342, DOI:10.1002/jez.2169.
- L. J. Legendre, S. Choi and J. A. Clarke, The diverse terminology of reptile eggshell microstructure and its effect on phylogenetic comparative analyses, J. Anat., 2022, 241(3), 641–666 CrossRef CAS PubMed.
- W. M. Saidel, N. Saglam, D. Salas-de la Cruz, R. Saunders and D. H. Shain, Elaborate ultrastructure of the Hirudo (Annelida: Hirudinae) cocoon surface, J. Morphol., 2018, 279(4), 545–553 CrossRef CAS PubMed.
- J. M. Garcia-Ruiz and A. B. Rodriguez-Navarro, The mineral structure of the avian eggshell: a case of competitive crystal growth, Bull. Inst. Oceanogr., 1994, 85–94 Search PubMed.
- A. Rodriguez-Navarro, C. Jimenez-Lopez, A. Hernandez-Hernandez, A. Checa and J. M. García-Ruiz, Nanocrystalline structures in calcium carbonate biominerals, J. Nanophotonics, 2008, 2(1), 021935 CrossRef.
- A. C. Fraser, M. M. Bain and S. E. Solomon, Organic matrix morphology and distribution in the palisade layer of eggshells sampled at selected periods during lay, Br. Poult. Sci., 1998, 39(2), 225–228 CrossRef CAS PubMed.
- M. Pines, V. Knopov and A. Bar, Involvement of osteopontin in egg shell formation in the laying chicken, Matrix Biol., 1995, 14(9), 765–771 CrossRef CAS PubMed.
- J. Gautron, M. T. Hincke, M. Panheleux, J. M. Garcia-Ruiz, T. Boldicke and Y. Nys, Ovotransferrin is a matrix protein of the hen eggshell membranes and basal calcified layer, Connect. Tissue Res., 2001, 42(4), 255–267 CrossRef CAS PubMed.
- S. Mandera, I. Coronado and L. Fernández-Díaz, et al., Earthworm granules: A model of non-classical biogenic calcium carbonate phase transformations, Acta Biomater., 2023, 162, 149–163 CrossRef CAS PubMed.
- Z. Zhang, Y. Xie, X. Xu, H. Pan and R. Tang, Transformation of amorphous calcium carbonate into aragonite, J. Cryst. Growth, 2012, 343(1), 62–67 CrossRef CAS.
- J. D. Rodriguez-Blanco, S. Shaw and L. G. Benning, The kinetics and mechanisms of amorphous calcium carbonate (ACC) crystallization to calcite, via vaterite, Nanoscale, 2011, 3(1), 265–271 RSC.
- J. Gautron, L. Stapane, N. Le Roy, Y. Nys, A. B. Rodriguez-Navarro and M. T. Hincke, Avian eggshell biomineralization: an update on its structure, mineralogy and protein tool kit, BMC Mol. Cell Biol., 2021, 22, 11 CrossRef CAS PubMed.
- M. T. Hincke, J. Gautron, M. Panheleux, J. Garcia-Ruiz, M. D. McKee and Y. Nys, Identification and localization of lysozyme as a component of eggshell membranes and eggshell matrix, Matrix Biol., 2000, 19(5), 443–453 CrossRef CAS PubMed.
- M. T. Hincke, Y. Nys and J. Gautron, The role of matrix proteins in eggshell formation, J. Poultry Sci., 2010, 47(3), 208–219 CrossRef CAS.
- M. L. H. Rose and M. T. Hincke, Protein constituents of the eggshell: eggshell-specific matrix proteins, Cell. Mol. Life Sci., 2009, 66, 2707–2719 CrossRef CAS PubMed.
- D. A. Carrino, J. E. Dennis and T. M. Wu, et al., The avian eggshell extracellular matrix as a model for biomineralization, Connect. Tissue Res., 1996, 35(1–4), 325–328 CrossRef CAS PubMed.
- N. Le Roy, L. Stapane, J. Gautron and M. T. Hincke, Evolution of the avian eggshell biomineralization protein toolkit–new insights from multi-omics, Front. Genet., 2021, 12, 672433 CrossRef CAS PubMed.
- A. V. Lensink, G. E. Swan and J. G. Myburg, The structure of the eggshell and eggshell membranes of Crocodylus niloticus, J. Microsc., 2023, 290(1), 23–39 CrossRef CAS PubMed.
- N. R. Elejalde-Cadena, D. Hernández and F. Capitelli, et al., Influence of Intramineral Proteins on the Growth of Carbonate Crystals Using as a Scaffold Membranes of Ratite Birds and Crocodiles Eggshells, Membranes, 2023, 13(11), 869 CrossRef CAS PubMed.
- J. Deering, V. Nelea and M. D. McKee, Multiscale Mineralization in the Leopard Gecko Eggshell, Adv. Funct. Mater., 2024, 34, 2316422 CrossRef CAS.
- N. R. Elejalde-Cadena, E. Hernández-Juárez, E. Tapia-Mendoza, A. Moreno and L. Bucio, Structural Insights into Ratite Birds and Crocodile Eggshells for Advanced Biomaterials Design, ACS Omega, 2025, 10, 5064–5079 CrossRef CAS PubMed.
- S. Choi, M. E. Hauber, L. J. Legendre, N. H. Kim, Y. N. Lee and D. J. Varricchio, Microstructural and crystallographic evolution of palaeognath (Aves) eggshells, eLife, 2023, 12, e81092 CrossRef CAS PubMed.
- V. R. Meenakshi, P. L. Blackwelder and N. Watabe, Studies on the formation of calcified egg-capsules of ampullarid snails, Calcif. Tissue Res., 1974, 16(1), 283–291, DOI:10.1007/BF02008236.
- R. H. Hackman and M. Goldberg, Composition of the oothecae of three Orthoptera, J. Insect Physiol., 1960, 5(1), 73–78 CrossRef CAS.
- M. Mazzini, M. Carcupino and A. M. Fausto, Egg chorion architecture in stick insects (Phasmatodea), Int. J. Insect Morphol. Embryol., 1993, 22(2–4), 391–415 CrossRef.
- T. van de Kamp and H. Greven, Comparative histological and scanning electron microscopical studies on the chorion of six species of stick insects (Phasmatodea), Acta Biol. Benrodis, 2016, 18, 45–127 Search PubMed.
- T. van de Kamp and H. Greven, Structure of the specialised and unspecialised chorion of the egg in the stick insect Malacomorpha cyllarum (Phasmatodea), Entomol. Gen., 2008, 31(1), 63–74 Search PubMed.
- K. E. Mikhailov, Fossil and recent eggshell in amniotic vertebrates: fine structure, comparative morphology and classification, Special Papers in Palaeontology, 1997, 56, 1–76 Search PubMed.
- M. W. J. Ferguson, The structure and composition of the eggshell and embryonic membranes of Alligator mississippiensis, Trans. Zool. Soc. Lond., 1982, 36(2), 99–152 CrossRef.
- S. Choi, S. Han, N. H. Kim and Y. N. Lee, A comparative study of eggshells of Gekkota with morphological, chemical compositional and crystallographic approaches and its evolutionary implications, PLoS One, 2018, 13(6), e0199496 CrossRef PubMed.
- H. J. Wu, Y. C. Tseng and S. H. Tsao, et al., A comparative study on the microstructures, mineral content, and mechanical properties of non-avian reptilian eggshells, Biology, 2023, 12(5), 688 CrossRef CAS PubMed.
- N. Le Roy, L. Combes-Soia and A. Brionne, et al., Guinea fowl eggshell quantitative proteomics yield new findings related to its unique structural characteristics and superior mechanical properties, J. Proteomics, 2019, 209, 103511 CrossRef CAS PubMed.
- M. Rousseau, A. Meibom, M. Gèze, X. Bourrat, M. Angellier and E. Lopez, Dynamics of sheet nacre formation in bivalves, J. Struct. Biol., 2009, 165(3), 190–195 CrossRef CAS PubMed.
- G. Debruyn, T. H. Büscher and M. P. J. Nicolaï, et al., Thin-film iridescence in the eggshell of a stick insect (Myronides glaucus), Physiol. Entomol., 2025, 50, 88–95 CrossRef CAS.
- K. F. Hirsch, Contemporary and fossil chelonian eggshells, Copeia, 1983, 382–397 CrossRef.
- S. Choi, H. Kim, I. Paik, Y. Park, H. Jung and X. Xu, Turtle eggs from the Lower Cretaceous Hasandong Formation (South Korea) with relict aragonite under significant thermal maturity, J. Vertebr. Paleontol., 2022, 42(4), e2183866 CrossRef.
- G. Krampitz and G. Graser, Molecular mechanisms of biomineralization in the formation of calcified shells, Angew. Chem., Int. Ed. Engl., 1988, 27(9), 1145–1156 CrossRef.
- R. Zhang, L. Xie and Z. Yan, Biomineralization Mechanism of the Pearl Oyster, Pinctada Fucata, Springer, 2018 Search PubMed.
- S. Kusuda, Y. Yasukawa and H. Shibata, et al., Diversity in the matrix structure of eggshells in the Testudines (Reptilia), Zool. Sci., 2013, 30(5), 366–374 CrossRef PubMed.
- A. C. Fraser, M. M. Bain and S. E. Solomon, Transmission electron microscopy of the vertical crystal layer and cuticle of the eggshell of the domestic fowl, Br. Poult. Sci., 1999, 40(5), 626–631 CrossRef CAS PubMed.
- H. Schraer, Biological Calcification: Cellular and Molecular Aspects, Springer Science & Business Media, 2012 Search PubMed.
- K. Alvares, The role of acidic phosphoproteins in biomineralization, Connect. Tissue Res., 2014, 55(1), 34–40 CrossRef CAS PubMed.
- L. Kalmar, D. Homola, G. Varga and P. Tompa, Structural disorder in proteins brings order to crystal growth in biomineralization, Bone, 2012, 51(3), 528–534 CrossRef CAS PubMed.
- Y. Nys, M. T. Hincke, J. L. Arias, J. M. Garcia-Ruiz and S. E. Solomon, Avian eggshell mineralization, Poultry and Avian Biology Reviews, 1999, 10(3), 143–166 Search PubMed.
- V. R. Franceschi and P. A. Nakata, Calcium oxalate in plants: formation and function, Annu. Rev. Plant Biol., 2005, 56(1), 41–71 CrossRef CAS PubMed.
- S. R. Khan, M. S. Pearle and W. G. Robertson, et al., Kidney stones, Nat. Rev. Dis. Primers, 2016, 2(1), 16008 CrossRef PubMed.
- T. Baird and S. E. Solomon, Calcite and aragonite in the egg shell of Chelonia mydas L., J. Exp. Mar. Biol. Ecol., 1979, 36(3), 295–303 CrossRef CAS.
- D. Konopacka-Łyskawa, Synthesis methods and favorable conditions for spherical vaterite precipitation: A review, Crystals, 2019, 9(4), 223 CrossRef.
- J. Dunning, A. Patil and L. D’Alba, et al., How woodcocks produce the most brilliant white plumagepatches among the birds, J. R. Soc. Interface, 2023, 20(200), 20220920 CrossRef PubMed.
- B. Vanthournout, F. Janssens and G. Debruyn, et al., Rapid and reversible humidity-dependent colour change by water film formation in a scaled springtail, J. R. Soc. Interface, 2023, 20(207), 20230228 CrossRef PubMed.
- S. Choi, Y. Park and M. Moreno-Azanza, Protocol for electron backscatter diffraction (EBSD) analysis of fossil eggshells, J. Vertebr. Paleontol., 2023, 43(6), e2363210 CrossRef.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d5fd00028a |
| This journal is © The Royal Society of Chemistry 2025 |