
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Uncovering biosynthetic relationships between antifungal nonadrides and octadrides†
Kate M. J.
de Mattos-Shipley‡
*ab,
Catherine E.
Spencer‡
 a,
Claudio
Greco
a,
David M.
Heard
a,
Daniel E.
O'Flynn
a,
Trong T.
Dao
a,
Zhongshu
Song
a,
Nicholas P.
Mulholland
c,
Jason L.
Vincent
c,
Thomas J.
Simpson
a,
Russell J.
Cox
d,
Andrew M.
Bailey
*b and
Christine L.
Willis
*a
a,
Claudio
Greco
a,
David M.
Heard
a,
Daniel E.
O'Flynn
a,
Trong T.
Dao
a,
Zhongshu
Song
a,
Nicholas P.
Mulholland
c,
Jason L.
Vincent
c,
Thomas J.
Simpson
a,
Russell J.
Cox
d,
Andrew M.
Bailey
*b and
Christine L.
Willis
*a
aSchool of Chemistry, University of Bristol, Cantock's Close, Bristol, BS8 1TS, UK. E-mail: Chris.Willis@bristol.ac.uk; andy.bailey@bristol.ac.uk; kd4495@bristol.ac.uk
bSchool of Biological Sciences, University of Bristol, 24 Tyndall Avenue, Bristol, BS8 1TQ, UK
cSyngenta, Jealott's Hill International Research Centre, Bracknell, RG42 6EY, UK
dInstitute for Organic Chemistry and BMWZ, Leibniz University of Hannover, Schneiderberg 38, 30167, Hannover, Germany
First published on 7th October 2020
Abstract
Maleidrides are a class of bioactive secondary metabolites unique to filamentous fungi, which contain one or more maleic anhydrides fused to a 7-, 8- or 9- membered carbocycle (named heptadrides, octadrides and nonadrides respectively). Herein structural and biosynthetic studies on the antifungal octadride, zopfiellin, and nonadrides scytalidin, deoxyscytalidin and castaneiolide are described. A combination of genome sequencing, bioinformatic analyses, gene disruptions, biotransformations, isotopic feeding studies, NMR and X-ray crystallography revealed that they share a common biosynthetic pathway, diverging only after the nonadride deoxyscytalidin. 5-Hydroxylation of deoxyscytalidin occurs prior to ring contraction in the zopfiellin pathway of Diffractella curvata. In Scytalidium album, 6-hydroxylation – confirmed as being catalysed by the α-ketoglutarate dependent oxidoreductase ScyL2 – converts deoxyscytalidin to scytalidin, in the final step in the scytalidin pathway. Feeding scytalidin to a zopfiellin PKS knockout strain led to the production of the nonadride castaneiolide and two novel ring-open maleidrides.
Introduction
Fungal maleidrides are an important family of polyketide-derived secondary metabolites which exhibit a diversity of biological activities including as antifungal1,2 and herbicidal3,4 agents.5 They are characterised by a medium-sized alicyclic ring with one or two fused maleic anhydride moieties. The majority of the reported maleidrides are nonadrides assembled on a 9-membered ring core, and early examples include byssochlamic acid 1,6 heveadride 2,7 glauconic acid 3 and glaucanic acid 4 (Fig. 1).8,9 Later studies revealed further nonadrides such as the phomoidrides, e.g.5,10 castaneiolide 6,11 rubratoxins, e.g.7![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 12 and cornexistin 8,3,13 the octadride viburspiran 9 and more recently two heptadrides agnestadrides A 10 and B have been isolated from Byssochlamys fulva.14 In certain cases the relative and absolute stereochemistry of the maleidrides remain unknown, and various details of their biosynthesis are yet to be fully elucidated.
12 and cornexistin 8,3,13 the octadride viburspiran 9 and more recently two heptadrides agnestadrides A 10 and B have been isolated from Byssochlamys fulva.14 In certain cases the relative and absolute stereochemistry of the maleidrides remain unknown, and various details of their biosynthesis are yet to be fully elucidated.
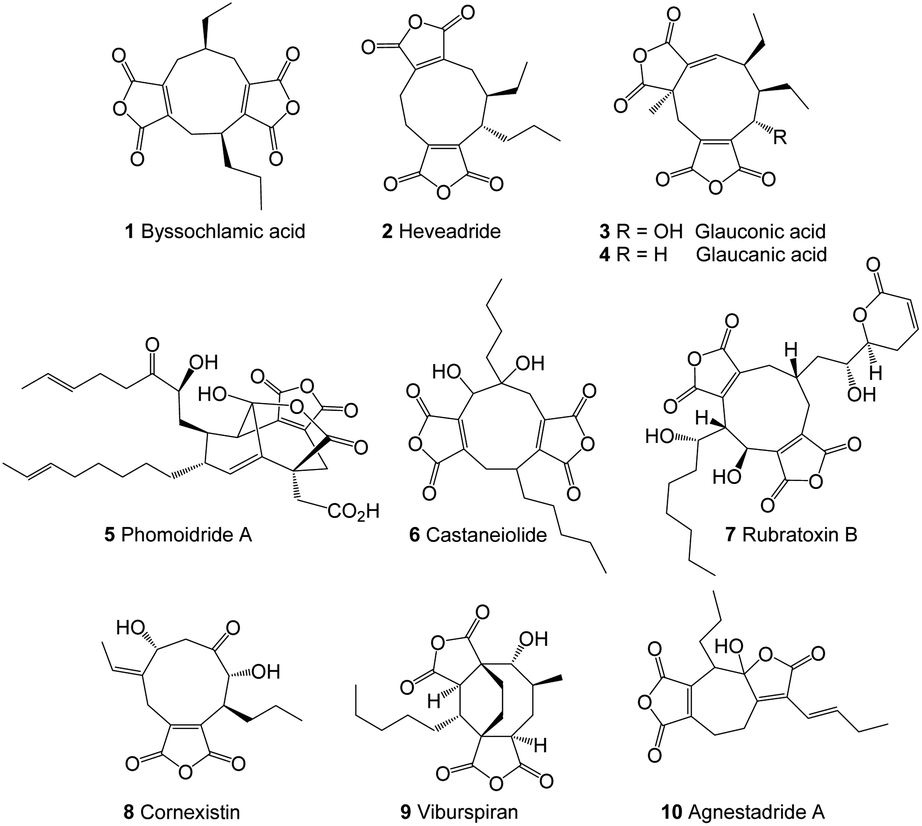 | ||
| Fig. 1 Examples of maleidrides. | ||
Results of extensive studies on selected nonadrides using isotopic labelling, gene knock outs and heterologous expression experiments have led to the proposal that the biosynthetic pathway begins with the assembly of an unsaturated precursor 11via an iterative highly reducing polyketide synthase (hrPKS), where the chain length varies according to the structure of the natural product (Scheme 1). Coupling of 11 with oxaloacetate is catalysed by citrate synthase-like enzymes and is followed by dehydration catalysed by 2-methylcitrate dehydratases to generate the maleidride monomer 12.15,16 Decarboxylation of 12 then gives the tautomeric compounds 13 and 14 (Scheme 1A).14,17 The second stage of the biosynthetic pathway, coupling the monomers in various modes and cyclisation to the carbocyclic ring, involves ketosteroid isomerase-like (KSI) and phosphatidylethanolamine binding protein-like (PEBP) enzymes.16 The mode of cyclisation determines the carbon framework of the maleidride (Scheme 1B), and finally tailoring modifications decorate these structures leading to the observed diversity of natural products.
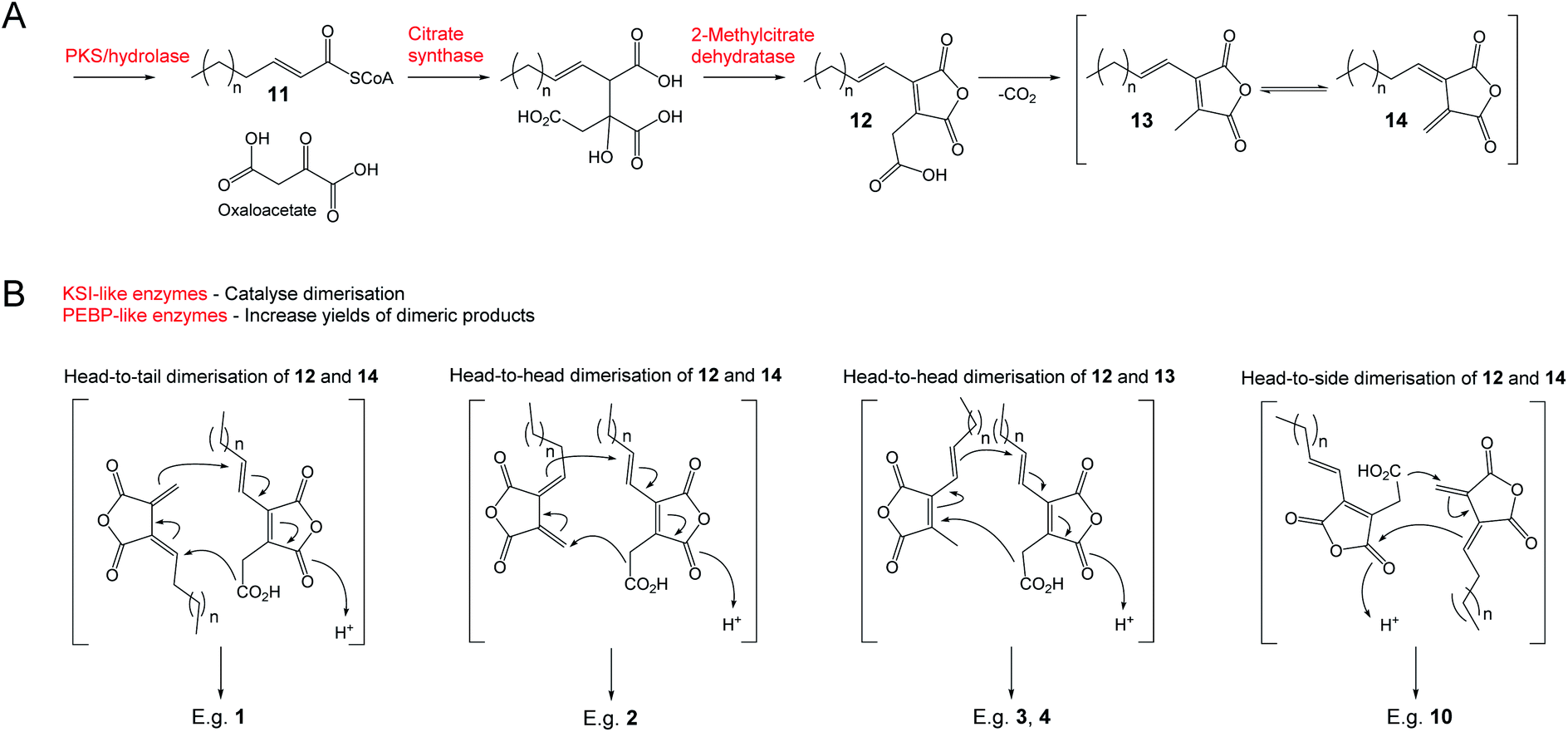 | ||
| Scheme 1 The universal pathway to maleidrides16 involves the production of maleic anhydride monomers (A) followed by dimerisation (B) – the mode of which determines the carbon framework of the final compound. | ||
Octadrides, with 8-membered rings, are less common than nonadrides and include viburspiran 918 and zopfiellin 1519 (Fig. 1 and 2). The biosynthetic origin of the 8-membered ring is unknown (§see footnote). Zopfiellin was first isolated from Zopfiella curvata and the structure determined by a combination of spectroscopic methods.19–21 The relative and absolute configurations of zopfiellin have been indicated in several publications,22–24 but we were unable to find any justification of the configuration of the side chain secondary alcohol. Hence, with our continuing interest in the biosynthesis of fungal natural products, and in particular bioactive maleidrides, our goal was to confirm the structure of zopfiellin 15 and determine the biosynthetic origin of the 8-membered ring, with the longer-term goal of engineering the pathway to new antifungal targets.
 | ||
| Fig. 2 Confirmed structures of the natural products 15, 16 and 17. | ||
Whilst this manuscript was in preparation Minami, Oikawa and co-workers reported investigations into the biosynthesis of zopfiellin using isotopic labelling, heterologous expression of candidate genes in Aspergillus oryzae, combined with structure elucidation of minor metabolites. These studies showed that a ring contraction occurs from a nonadride to an octadride.25 Herein we describe our recent studies which confirm the absolute and relative configurations of the nonadride scytalidin 16 and octadride zopfiellin 15 and reveal that they share a common biosynthetic precursor, deoxyscytalidin 17 (Fig. 2). Our biosynthetic investigations involved a different approach from those previously reported25 and used a combination of gene disruptions in Diffractella curvata and Scytalidium album, feeding studies to mutant strains and full structure elucidation of known and novel compounds. These studies establish the biosynthetic relationships between zopfiellin 15, scytalidin 16, and deoxyscytalidin 17.
Results and discussion
The confirmed zopfiellin producer, Zopfiella curvata no. 37-3, was not publicly available when these studies began, so Diffractella curvata CBS591.74 was obtained and confirmed to produce zopfiellin. Cultures were grown in shake flasks in PDB medium at 25 °C for 14 days and after purification by flash chromatography, gave zopfiellin 15 (80–100 mg L−1) as an oil. As full assignment of NMR data had not previously been reported19,21 2D NMR data (COSY, HSQC and HMBC) analysis allowed assignment of the 1H and 13C NMR spectra (Table S6 and Fig. S31–S36†). The optical rotation, [α]D −84.3 (c 0.43 MeOH) was consistent with the literature value, [α]D −76.8 (c 0.42 MeOH).20 Zopfiellin was esterified to give the novel p-nitrobenzoate 18 and X-ray crystallography revealed the relative configuration of the natural product (Fig. 2). Synthesis and analysis of the (R)- and (S)-Mosher's ester derivatives of 15 confirmed the absolute stereochemistry of zopfiellin as 1R, 5R, 1′′R (Fig. S81–S83†).Feeding [1,2-13C2]-acetate to cultures of D. curvata showed mainly intact incorporation of acetate into zopfiellin 15 (Fig. 3). Enhancement of C-5 in the 13C NMR spectrum was observed, but there was no coupling to an adjacent 13C, indicating that it originates from a cleaved acetate and therefore that a carbon atom is lost during zopfiellin biosynthesis.
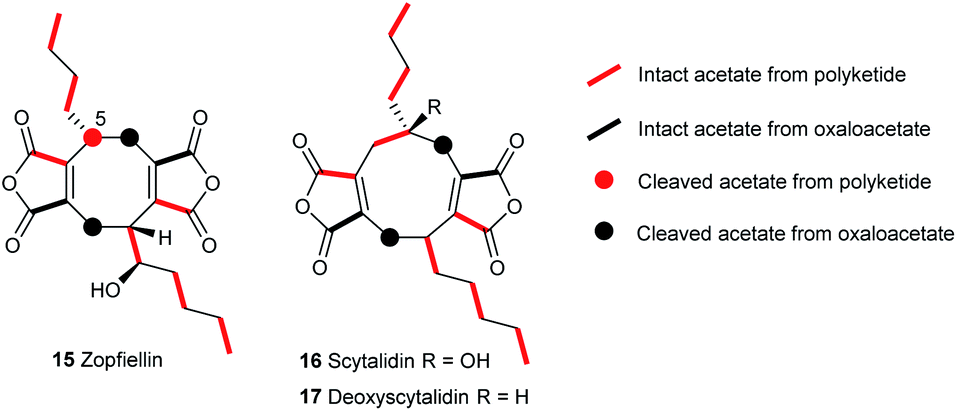 | ||
| Fig. 3 Incorporation patterns in octadrides and nonadrides from feeding studies with [1,2-13C2]-acetate. | ||
A possible biosynthetic precursor to zopfiellin is therefore a nonadride which on rearrangement with the loss of one carbon atom would give the corresponding octadride. Such ring contractions are well precedented in fungal biosynthesis, for example, in the contraction of ring B of the kaurenoids to the gibberellins,26 the conversion of the 14-membered polyketide macrodiolide colletotriene to the 13-membered ring of bartanol27 and the oxidative ring contractions seen in xenovulene biosynthesis.28 Conceivably the known natural products scytalidin 1629 (C22H28O7), deoxyscytalidin 1730 (C22H28O6) or castaneiolide 611 (C22H28O8) could be precursors. To obtain samples of 16 and 17, cultures of S. album strain UAMH 3620 were grown, and after 14 days compounds 16 and 17 were isolated in similar titres (ca. 10 mg L−1). X-ray analysis confirmed that the alkyl side chains are syn in both metabolites (Fig. 2). The results of feeding studies using [1,2-13C2] acetate to cultures of S. album were in accord with the predicted biosynthetic pathway to nonadrides and that one or both could possibly be biosynthetic precursors of zopfiellin 15 (Fig. 3).
De novo genome sequencing was performed for D. curvata CBS591.74 and S. album UAMH3620, and an initial screen of the genome data, searching for homologues to the byssochlamic acid biosynthetic gene cluster (BGC),16 revealed putative maleidride gene clusters within both genomes. The gene clusters were analysed in detail using FGeneSH31 to predict coding sequences, and manually annotated in Artemis32 to produce full draft BGCs (Fig. 4 and Table 1). Putative gene functions were assigned via the identification of homologues in the NCBI database and the detection of conserved domains using InterPro33 (Tables S1 and S2†). Transcriptomic analysis for D. curvata under a range of zopfiellin 15 production and non-production conditions aided identification of the cluster boundaries by clearly identifying a co-transcribed region that includes genes from zopL9 to zopR4 (Fig. 4, S1 and S2†). In addition, detailed analysis of the transcriptomic data confirmed the positions of all introns and exons in the predicted ORFs. However, despite repeated attempts, it was not possible to generate quality RNAseq data for the putative scytalidin gene cluster in S. album.
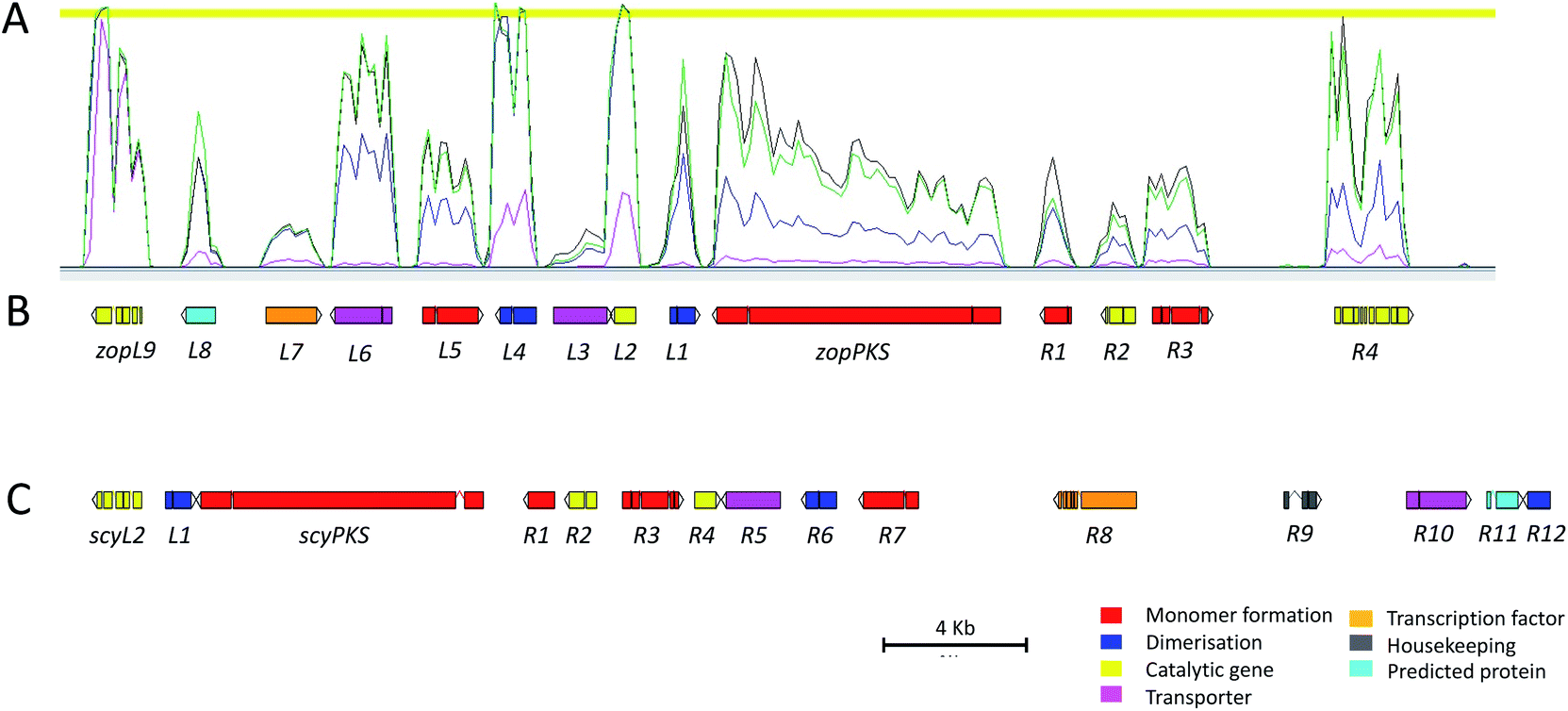 | ||
| Fig. 4 (A) RNAseq data mapped to the zopfiellin BGC showing coregulation of the genes under four different conditions (Fig. S1 and S2†). Pink: GN. Purple: CDB. Green: PDB-day 5. Black: PDB-day 8. (B) The zopfiellin gene cluster, as annotated in Artemis, which contains genes zopL9 to zopR4. (C) The scytalidin gene cluster, as annotated in Artemis, which contains genes scyL2 to scyR12. ‘L’ in the gene names denotes genes to the left, or upstream, of the PKS. ‘R’ denotes genes to the right, or downstream, of the PKS. Putative gene functions are shown in Table 1. | ||
| Gene | Putative function | Homologue | Gene | Putative function | Homologue |
|---|---|---|---|---|---|
| zopL9 | Hydroxylase/Desaturase | DES34 | scyL2 | Hydroxylase | AclN36 |
| AsaB35 | MfR137 |
||||
| zopL8 | Hypothetical protein | — | scyL1* | PEBP | BfL516 |
| zopL7 | Transcription factor (TF) | AlnR38 | scyPKS | hrPKS | Tox1-PKS39 |
| zopL6 | Major facilitator superfamily (MFS) transporter | Itp140 |
scyR1 | DUF341 hydrolase/esterase | Fub441 |
| zopL5 | 2-Methylcitrate dehydratase (2MCDH) | PrpD42 | scyR2 | Enoyl CoA hydratase | Ech143 |
| zopL4* | Ketosteroid isomerase-like protein (KSI) | BfL616 |
scyR3 | Citrate synthase | MfR344 |
| zopL3 | Major facilitator superfamily (MFS) transporter | Itp140 |
scyR4 | Isochorismatase-like hydrolase/amidohydrolase | CSHase45 |
| zopL2 | Isochorismatase-like hydrolase/amidohydrolase | NicF46 | scyR5 | MFS transporter | MfM644 |
| scyR6* | KSI | BfL616 |
|||
| zopL1 | Phosphatidylethanolamine-binding protein (PEBP) | Tfl147 |
scyR7 | 2-Methylcitrate dehydratase | PrpD42 |
| zopPKS | hrPKS | Tox1-PKS39 | scyR8 | Transcription factor | AlnR38 |
| zopR1 | DUF341 hydrolase/esterase | Fub441 |
scyR9 | Histone H2A | H2A.Z48 |
| zopR2 | Enoyl-CoA hydratase | Ech143 |
scyR10 | MFS transporter | MfR544 |
| zopR3 | Citrate synthase | CshA49 | scyR11 | Hypothetical protein | — |
| zopR4 | FAD-dependent oxidoreductase | YanF50 | scyR12* | PEBP | BfL916 |
| Sol551,52 |
To confirm the identity of the two gene clusters, protoplast-mediated transformation protocols were developed for both fungal species and the PKS genes; zopPKS and scyPKS, were disrupted using the bipartite gene knock-out approach developed by Nielsen et al.53 In both cases this led to a total loss of maleidride biosynthesis, with no scytalidin 16 or deoxyscytalidin 17 being present in the crude extracts of the ΔscyPKS strains (Fig. 5e), and no zopfiellin 15 being present in the crude extracts of the ΔzopPKS strains (Fig. 5h).
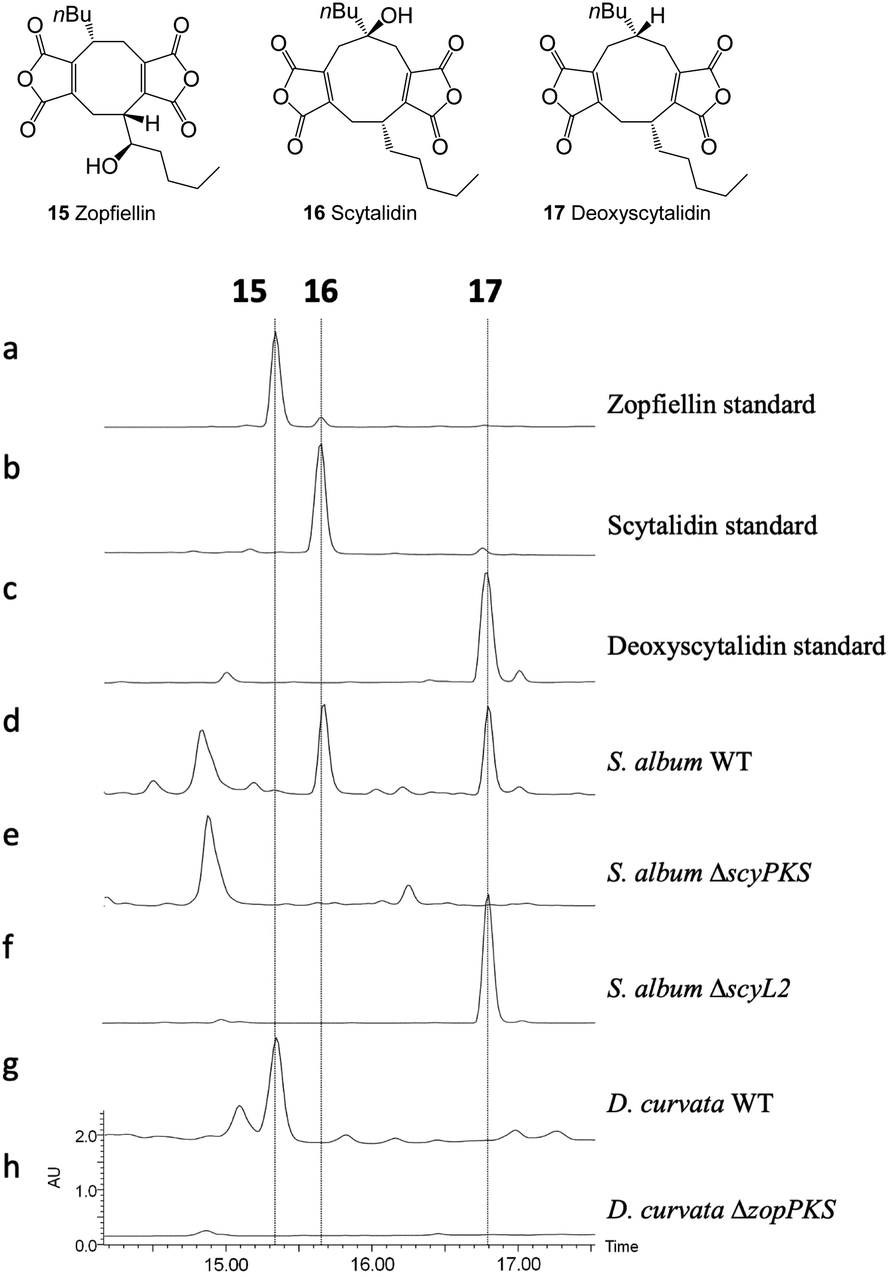 | ||
| Fig. 5 HPLC (DAD) analysis of gene deletion strains for the zopfiellin and scytalidin gene clusters. Disruption of zopPKS and scyPKS abolished maleidride production in both species, confirming the identity of the zopfiellin and scytalidin BGCs. Disruption of scyL2 confirmed its role in the final hydroxylation of the scytalidin pathway. | ||
An ACT (Artemis Comparison Tool)54 comparison of the zopfiellin and scytalidin BGCs (Fig. 6) highlighted significant homologies. Both clusters contain genes encoding the essential hrPKS, DUF341 hydrolase, citrate synthase (CS) and 2-methylcitrate dehydratase (2MCDH),12,13,15,16 as well as the ‘dimerisation’ genes encoding a ketosteroid-isomerase-like protein (KSI) and a PEBP-like protein.16 In the case of the scytalidin BGC a second PEBP gene is present, which has been observed previously in the byssochlamic acid and rubratoxin BGCs.12,16 A domain analysis of the two PKS genes identified all of the domains typically present in a highly-reducing PKS: KS, AT, DH, CMeT, ER, KR and ACP. Additional genes identified in both clusters encode an enoyl-CoA hydratase, an isochorismatase, a small hypothetical protein (zopL8 and scyR11), a GAL4 type transcription factor and two MFS (major facilitator superfamily) transporters. The enoyl-CoA hydratase encoded by the zopfiellin cluster (zopR2) was not identified in the work by Oikawa and colleagues,25 but RNAseq data confirmed its annotation.
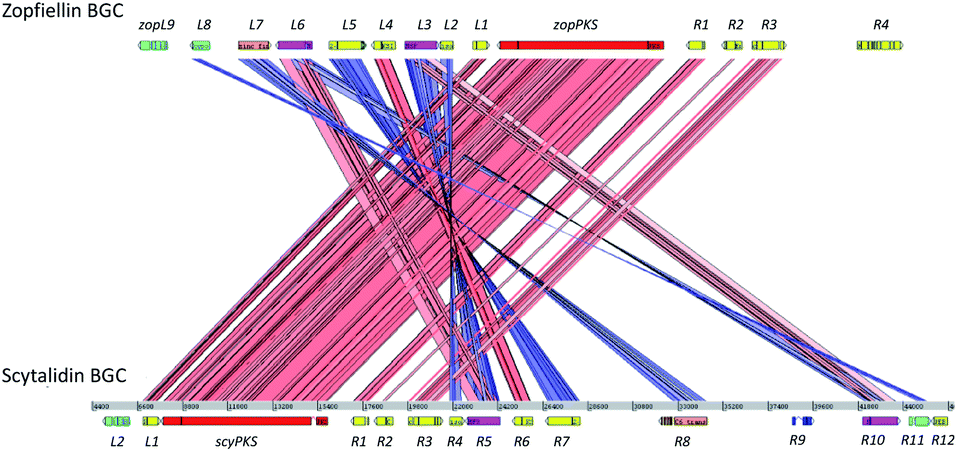 | ||
| Fig. 6 An ACT (Artemis Comparison Tool) comparison of the zopfiellin and scytalidin BGCs identified genes predicted to encode homologous proteins and highlighted the similarity between the two biosynthetic gene clusters. | ||
The proteins encoded by the clusters also have high sequence identity (Table S3†). The KSI and PEBP-like proteins, for example, are far more similar to one another (80.7% and 66.2% identity respectively, Table S3†) than to those identified in other maleidride BGCs (Fig. S4–S7†). In the case of the transporters, ZopL6 and ScyR5 are highly homologous (80.4% identity) and thus may be expected to transport similar compounds. The other two transporters, ZopL3 and ScyR10, are more diverse, sharing only 38.5% identity. All other homologues encoded by the gene clusters share over 50% identity, with many sharing over 80% (Table S3†).
The only proteins which are uniquely encoded by the zopfiellin gene cluster are a putative hydroxylase/desaturase (ZopL9), which shares homology with DES from the gibberellin pathway, and an FAD-dependent oxidoreductase (ZopR4). The scytalidin gene cluster encodes only one unique protein, a putative hydroxylase (ScyL2). ZopL9 and ScyL2 are both predicted to be α-ketoglutarate (αKG) dependent enzymes based on the presence of conserved iron and αKG binding residues (Fig. S10†), but they were not considered to be homologues due to the low level of sequence identity (approximately 25% identity), suggesting that the exact catalytic function is unlikely to be the same. Also located within the scytalidin BGC is a gene (scyR9) encoding a highly conserved H2A.F/Z family histone,55 but this is unlikely to play a direct role in scytalidin biosynthesis (Table S3†).
The striking similarity between the two gene clusters (Fig. 6) suggests that zopfiellin 15 and scytalidin 16 have very similar biosynthetic origins, and that either deoxyscytalidin 17 or scytalidin 16 may be precursors of zopfiellin. To test this theory, 16 and 17 were purified from S. album cultures and separately fed to D. curvata ΔzopPKS cultures on days 3 and 5 post-inoculation (5 mg per day per 100 ml of culture). In cultures fed with 17, zopfiellin production was restored (Scheme 2 and Fig. 7e), demonstrating that deoxyscytalidin 17 is a precursor of zopfiellin. The extent of conversion of deoxyscytalidin to zopfiellin varied between individual feeds, presumably due to varying expression levels for the zopfiellin BGC. In cultures where the conversion was partial, the HPLC trace showed a peak for deoxyscytalidin, as well as a novel compound with a mass of 404 eluting at 16.1 minutes (Fig. 7e). This metabolite was isolated, and the structure confirmed by NMR spectroscopy to be 5-hydroxy-deoxyscytalidin 19 (Table S9 and Fig. S44–S49†). The signal assigned to 5-H (at δ 4.90) appeared as a broad singlet and NOE studies were in accord with hydroxylation occurring on the same face as the butyl side chain at C-6. To determine whether 19 is a shunt product or an intermediate on the biosynthetic pathway, it was fed to D. curvata ΔzopPKS cultures and again this restored zopfiellin biosynthesis (Scheme 2 and Fig. 7f).
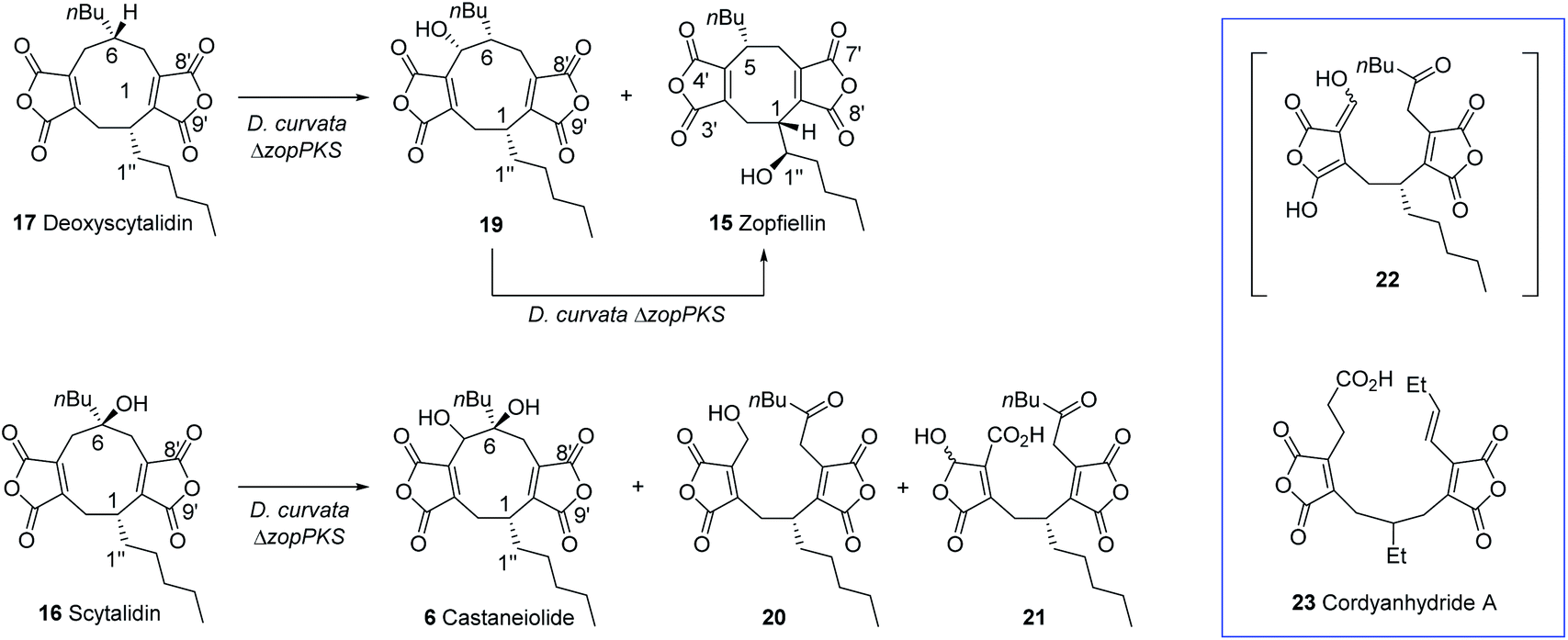 | ||
| Scheme 2 Feeding studies to D. curvata strain ΔzopPKS identified 17 and 19 as intermediates in zopfiellin biosynthesis and scytalidin was metabolised to novel products. | ||
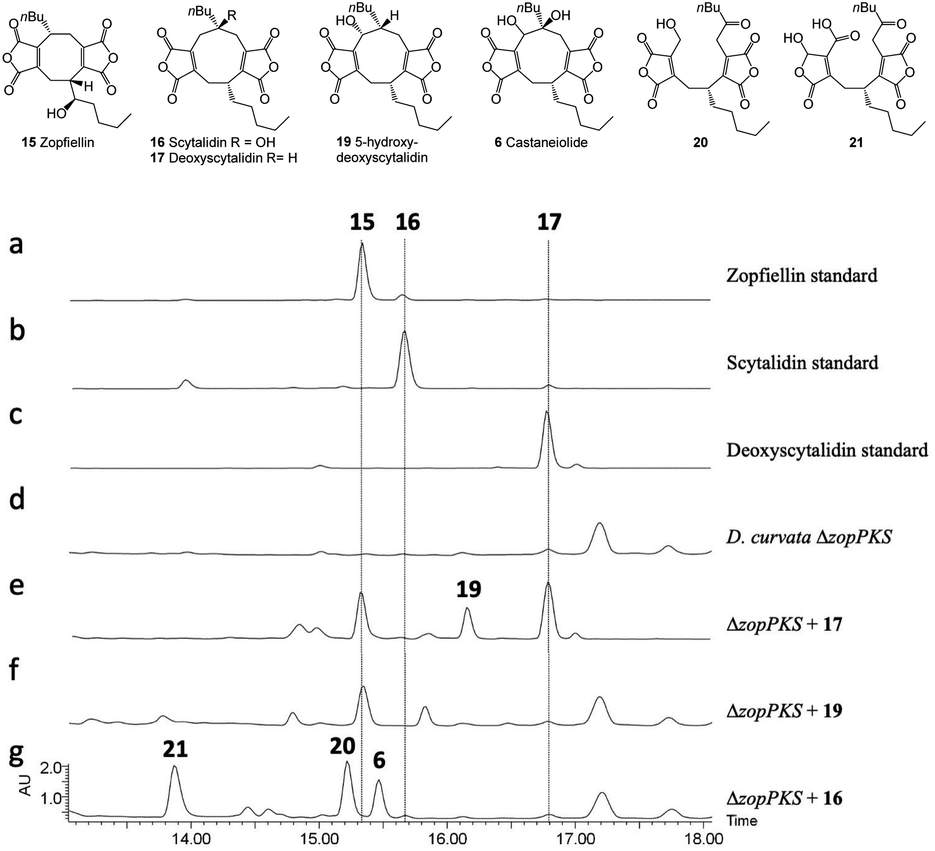 | ||
| Fig. 7 HPLC (DAD) chromatograms of crude extracts. Feeding compounds to D. curvata ΔzopPKS indicate that deoxyscytalidin 17 and 5-hydroxy-deoxyscytalidin 19 are intermediates in the zopfiellin biosynthetic pathway, whereas scytalidin 16 is not. Feeding of 16 led to the production of castaneiolide 6 and two novel ring-open anhydrides 20 and 21. | ||
When scytalidin 16 was fed to cultures of D. curvata ΔzopPKS, no zopfiellin was detected, but several additional compounds were present when compared to controls (Fig. 7g). The mixture was purified by HPLC giving the known11 nonadride castaneiolide 6 as one of the metabolites, presumably formed by 5-hydroxylation of scytalidin 16 (Scheme 2). In addition, two novel compounds 20 and 21 were isolated (3.2 mg and 0.7 mg respectively) and their structures elucidated by MS and extensive NMR studies (Table S10 and Fig. S50–S63†). The 13C NMR spectrum of compound 20 (C22H28O8 from HRMS) showed downfield signals assigned to the two maleic anhydrides, as well as an additional signal at δC 205 ppm assigned to a ketone. The 1H-NMR showed characteristic AB signals assigned to the hydroxymethylene. Based on this evidence, and combined with further analysis of 2D NMR data, the new metabolite was assigned as the ring cleaved alcohol 20. The spectral data of 21 was also in accord with a ring cleaved metabolite with a ketone in the side chain, but in this case the primary alcohol was oxidised to a carboxylic acid and one of the carbonyls of the anhydride reduced to give a mixture of epimers (δC 96.9/97.4 ppm, δH 6.19/6.23 ppm). Such reductions of anhydrides are commonly observed in maleidride biosynthesis.14,56,57 We propose that ketone 20 may be formed via cleavage of the 5,6-diol 6 to give an intermediate 22, which would tautomerise to generate ketone 20. Oxidation of the primary alcohol of 20 to a carboxylic acid and selective reduction of the anhydride would give 21. Such linear bis(maleic anhydride) structures are not without precedent, for example the fungal metabolite cordyanhydride A 23.58 However, 23 has previously been proposed to arise via the linear coupling of two distinct maleic anhydride monomers rather than via cleavage of a cyclic intermediate.15
Based on the bioinformatic analysis of the BGCs and the identification of deoxyscytalidin 17 as an intermediate to both scytalidin 16 and zopfiellin 15, it is a reasonable inference that the two biosynthetic pathways proceed in an identical manner to 17, after which they diverge. Hydroxylation at C-6 would produce scytalidin whereas hydroxylation at C-5, ring contraction and hydroxylation of the side chain would produce zopfiellin.
ScyL2, being the only unique catalytic protein encoded by the scytalidin BGC, was identified as a likely candidate for catalysing the final step in the scytalidin biosynthetic pathway. This is supported by a bioinformatic analysis identifying ScyL2 as a likely α-ketoglutarate dependent hydroxylase (Table S2†), which shares homology with enzymes known to catalyse hydroxylations, such as MfR1 and MfR2 from the squalestatin pathway,37 and RbtB and RbtG from the rubratoxin pathway12 (Fig. S8–S10†). To confirm this role, scyL2 knock-out strains of S. album were generated. These strains did not produce scytalidin 16, but accumulated deoxyscytalidin 17 (Fig. 5f), confirming the role of ScyL2.
Recent work by Shiina et al.25 on the zopfiellin pathway of Z. curvata, using heterologous expression as well as in vitro assays, has begun to elucidate the later stages of zopfiellin biosynthesis. ZopK, which is equivalent to ZopL9 of the D. curvata BGC reported here, was shown to catalyse the oxidative ring contraction in the zopfiellin pathway to produce deoxyzopfiellin 24.25 This proceeds via the intermediate 19 (which we now know to be 5-hydroxy-deoxyscytalidin) and is consistent with our identification, via feeding studies, of 19 as an intermediate in the zopfiellin pathway. In vitro assays using purified recombinant ZopL9 gave results which are entirely consistent with the findings of Shiina et al.25 In the presence of α-ketoglutarate, ZopL9 was capable of catalysing highly efficient hydroxylation of deoxyscytalidin 17 to give 5-hydroxy-deoxyscytalidin 19 (Fig. S68†). A minor compound was also detected which was purified and identified as deoxyzopfiellin 24. Repeating the assays using 5-hydroxy-deoxyscytalidin 19 as the substrate also gave rise to deoxyzopfiellin 24 (Fig. S68†), in accord with the proposal that ZopL9 is implicated in the ring contraction in the zopfiellin pathway of D. curvata, by first hydroxylating deoxyscytalidin to give 19, then catalysing the ring contraction to give the octadride core.
Having identified 17, 19 and 24 as intermediates in the zopfiellin pathway – through both feeding studies and in vitro assays – an extract of wild-type D. curvata was re-analysed, which revealed the presence of all three compounds as minor metabolites (Fig. S25†). Taken together with the bioinformatic analysis and the discovery that ScyL2 catalyses the final step of the scytalidin pathway, a branching biosynthetic route for the production of deoxyscytalidin, scytalidin and zopfiellin can now be proposed as shown in Scheme 3.
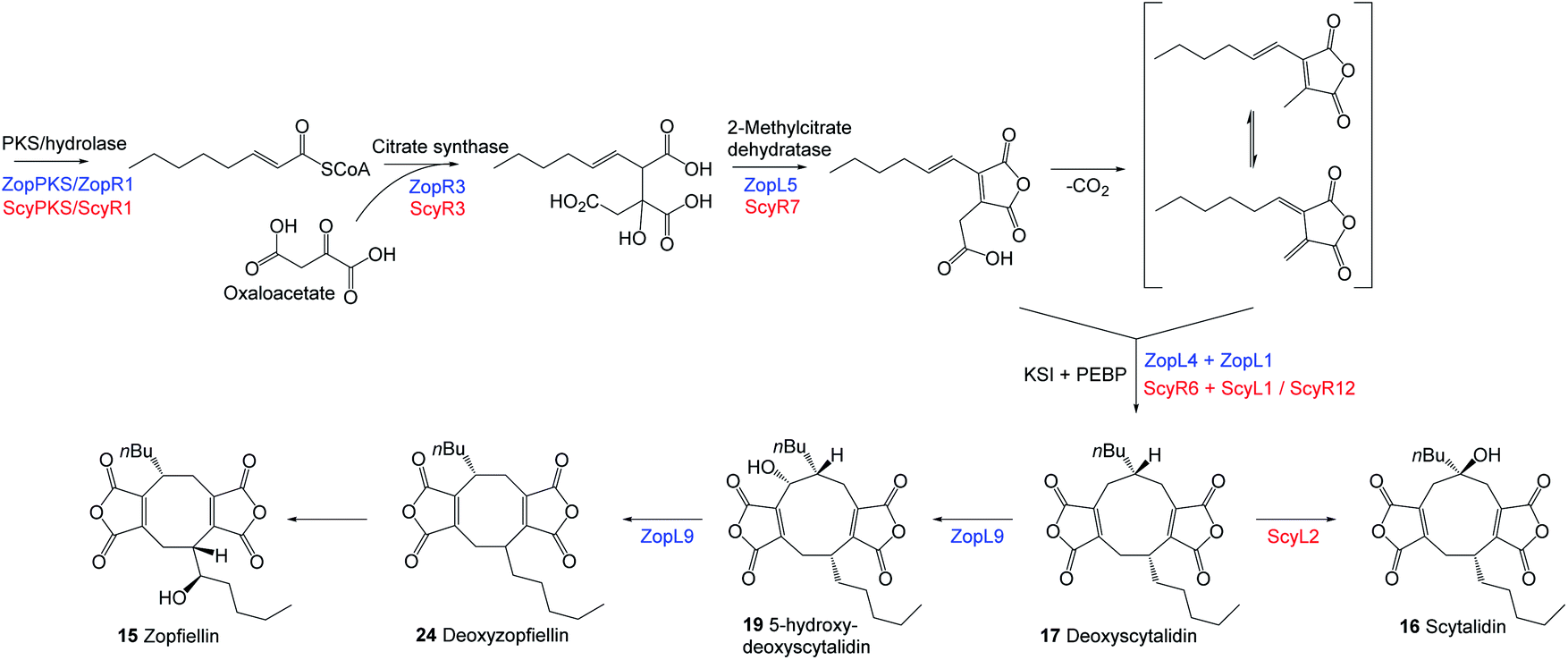 | ||
| Scheme 3 Proposed biosynthetic pathways of the maleidrides deoxyscytalidin 17, scytalidin 16 and zopfiellin 15. | ||
Intriguingly there is a homologue to ZopL9 (PhiK) encoded by the phomoidride BGC (44.2% homology – Fig. S11†). The structure of the phomoidrides, e.g.5, suggests that multiple oxidations must occur during their biosynthesis, but no ring contraction of the nine-membered core occurs in this pathway, and the role of PhiK has not yet been investigated.
Conclusions
This work categorically establishes the biosynthetic relationship between the nonadride scytalidin 16 and the octadride zopfiellin 15, via the shared nonadride intermediate; deoxyscytalidin 17. In both pathways, deoxyscytalidin is the substrate for αKG-dependent dioxygenase enzymes. In the scytalidin pathway ScyL2 catalyses a 6-hydroxylation of deoxyscytalidin to produce scytalidin whereas in the zopfiellin pathway ZopL9 catalyses a 5-hydroxylation of deoxyscytalidin and is implicated in a ring contraction to produce the octadride core. Interestingly, when cultures of D. curvata ΔzopPKS were fed with scytalidin, (possessing a 6-hydroxy group), 5-hydroxylation occurred to give castaneiolide 6 and two novel ring-cleaved maleidrides 20 and 21 were produced. Genes encoding αKG-dependent enzymes have been identified within a further three maleidride BGCs; namely rbtB, rbtE, rbtG and rbtU of the rubratoxin cluster, and the currently uncharacterised phiK of the phomoidride BGC and pvL5 of the cornexistin BGC. This highlights the important role these, often multifunctional, enzymes play in generating the structural diversity seen in the maleidride class of natural products. Intriguing questions which remain to be answered regarding the biosynthesis of zopfiellin include the mechanism by which the ring contraction occurs, and the identification of the enzyme responsible for installing the hydroxyl group at C-1′′ of the side chain of zopfiellin. ZopR4, which is uniquely encoded by the zopfiellin BGC and predicted to be a FAD-dependent oxidoreductase, is a candidate for catalysing such a transformation, but this remains to be explored experimentally. Additionally, the roles of the isochorismatase-like enzyme (ZopL2/ScyR4), the enoyl CoA hydratase (ZopR2/ScyR2) and the small hypothetical protein (ZopL8/ScyR11), which are encoded by both BGCs, are currently unknown.Conflicts of interest
There are no conflicts to declare.Acknowledgements
We thank BBSRC (BB/K002341/1) and Syngenta for funding (KMJdMS, CG and ZS) and a PhD studentship BB/P504804/1 (CS). We are also grateful to MRC (MR/N029909/1) (funding for TTD) and EPSRC (EP/L015366/1), Bristol Chemical Synthesis Centre for Doctoral Training, which provided PhD studentships (DMH and DOF). We are also very grateful to BBSRC and EPSRC for funding instrumentation via the Bristol Centre for Synthetic Biology (BB/L01386X/1). Genome sequencing, the production of assembled draft genomes and RNA sequencing were all carried out at the DNA sequencing facility in the Biochemistry Department of Cambridge University. X-ray crystallography was carried out by Dr Hazel Sparkes, School of Chemistry, University of Bristol. In additional we are grateful to Dr Lauren Ray from Syngenta for her help and support during the Industrial placement for CS.Notes and references
- T. Hosoe, K. Fukushima, T. Itabashi, K. Nozawa, K. Takizawa and K.-I. Kawai, Heterocycles, 2004, 63, 2581–2589 CrossRef CAS.
- M. Futagawa, A. M. Rimando, M. R. Tellez and D. E. Wedge, J. Agric. Food Chem., 2002, 50, 7007–7012 CrossRef CAS.
- M. Nakajima, K. Itoi, Y. Takamatsu, S. Sato, Y. Furukawa, K. Furuya, T. Honma, J. Kadotani, M. Kozasa and T. Haneishi, J. Antibiot., 1991, 44, 1065–1072 CrossRef CAS.
- S. C. Fields, L. Mireles-Lo and B. C. Gerwick, J. Nat. Prod., 1996, 59, 698–700 CrossRef CAS.
- R. Schor and R. Cox, Nat. Prod. Rep., 2018, 35, 230–256 RSC.
- H. Raistrick and G. Smith, Biochem. J., 1933, 27, 1814–1819 CrossRef CAS.
- R. I. Crane, P. Hedden, J. MacMillan and W. B. Turner, J. Chem. Soc., Perkin Trans. 1, 1973, 194–200 RSC.
- N. Wijkman, Justus Liebigs Ann. Chem., 1931, 485, 61–73 CrossRef CAS.
- J. E. Baldwin, D. H. R. Barton, J. L. Bloomer, L. M. Jackman, L. Rodriguez-Hahn and J. K. Sutherland, Experientia, 1962, 18, 345–352 CrossRef CAS.
- P. Spencer, F. Agnelli and G. A. Sulikowski, Org. Lett., 2001, 3, 1443–1445 CrossRef CAS.
- K. Arai, S. Shimizu, H. Miyajima and Y. Yamamoto, Chem. Pharm. Bull., 1989, 37, 2870–2872 CrossRef CAS.
- J. Bai, D. Yan, T. Zhang, Y. Guo, Y. Liu, Y. Zou, M. Tang, B. Liu, Q. Wu, S. Yu, Y. Tang and Y. Hu, Angew. Chem., Int. Ed., 2017, 56, 4782–4786 CrossRef CAS.
- K. Williams, A. J. Szwalbe, C. Dickson, T. R. Desson, N. P. Mulholland, J. L. Vincent, J. M. Clough, A. M. Bailey, C. P. Butts and C. L. Willis, Chem. Commun., 2017, 53, 7965–7968 RSC.
- A. J. Szwalbe, K. Williams, D. E. O'Flynn, A. M. Bailey, N. P. Mulholland, J. L. Vincent, C. L. Willis, R. J. Cox and T. J. Simpson, Chem. Commun., 2015, 51, 17088–17091 RSC.
- R. Fujii, Y. Matsu, A. Minami, S. Nagamine, I. Takeuchi, K. Gomi and H. Oikawa, Org. Lett., 2015, 17, 5658–5661 CrossRef CAS.
- K. Williams, A. J. Szwalbe, N. P. Mulholland, J. L. Vincent, A. M. Bailey, C. L. Willis, T. J. Simpson and R. J. Cox, Angew. Chem., Int. Ed., 2016, 55, 6784–6788 CrossRef CAS.
- D. M. Heard, E. R. Tayler, R. J. Cox, T. J. Simpson and C. L. Willis, Tetrahedron, 2020, 76, 130717 CrossRef CAS.
- M. Saleem, H. Hussain, I. Ahmed, S. Draeger, B. Schulz, K. Meier, M. Steinert, G. Pescitelli, T. Kurtan, U. Flörke and K. Krohn, Eur. J. Org Chem., 2011, 2011, 808–812 CrossRef.
- Y. Nakajima, H. Watanabe, T. Adachi, M. Tagawa, M. Futagawa and Y. Nishino, Japan Patent, JP08107792, 1996.
- T. Watanabe, T. Yasumoto, M. Murata, M. Tagawa, H. Narushima, T. Furusato, M. Kuwahara, M. Hanaue and T. Seki, Europe Patent, EP0582267A1, 1994.
- A. Nagamura, H. Aoki, T. Nishimura and I. Kinoshita, Japan Patent, JP09087268, 1997.
- Y. Baba, C. Noutary, S. Ichikawa, T. Kusumoto, H. Watanabe, M. Adachi, H. Hotta, Y. Nakajima, K. Arai and T. Hiyama, Synlett, 1997, 12, 1393–1394 CrossRef.
- H. Watanabe, Y. Nakajima, M. Adachi, H. Hotta, K. Arai, Y. Baba, C. Noutary, S. Ichikawa and T. Kusumoto, Chem. Commun., 1999, 17, 1753–1754 RSC.
- L. Musso, S. Dallavalle, G. Farina and E. Burrone, Chem. Biol. Drug Des., 2012, 79, 780–789 CrossRef CAS.
- T. Shiina, T. Ozaki, Y. Matsu, S. Nagamine, C. Liu, M. Hashimoto, A. Minami and H. Oikawa, Org. Lett., 2020, 22, 1997–2001 CrossRef CAS.
- B. E. Cross, K. Norton and J. C. Stewart, J. Chem. Soc. C, 1968, 1054–1063 RSC.
- K. Hanson, J. A. O’Neill, T. J. Simpson and C. L. Willis, J. Chem. Soc., Perkin Trans. 1, 1994, 17, 2493–2497 RSC.
- R. Schor, C. Schotte, D. Wibberg, J. Kalinowski and R. J. Cox, Nat. Commun., 2018, 9, 1963 CrossRef.
- G. M. Strunz, M. Kakushima and M. A. Stillwell, J. Chem. Soc., Perkin Trans. 1, 1972, 18, 2280–2283 RSC.
- W. A. Ayer, P. P. Lu, H. Orszanska and L. Sigler, J. Nat. Prod., 1993, 56, 1835–1838 CrossRef CAS.
- V. Solovyev, P. Kosarev, I. Seledsov and D. Vorobyev, Genome Biol., 2006, 7, S10 CrossRef.
- K. Rutherford, J. Parkhill, J. Crook, T. Horsnell, P. Rice, M.-A. Rajandream and B. Barrell, Bioinformatics, 2000, 16, 944–945 CrossRef CAS.
- R. D. Finn, T. K. Attwood, P. C. Babbitt, A. Bateman, P. Bork, A. J. Bridge, H.-Y. Chang, Z. Dosztányi, S. El-Gebali, M. Fraser, J. Gough, D. Haft, G. L. Holliday, H. Huang, X. Huang, I. Letunic, R. Lopez, S. Lu, A. Marchler-Bauer, H. Mi, J. Mistry, D. A. Natale, M. Necci, G. Nuka, C. A. Orengo, Y. Park, S. Pesseat, D. Piovesan, S. C. Potter, N. D. Rawlings, N. Redaschi, L. Richardson, C. Rivoire, A. Sangrador-Vegas, C. Sigrist, I. Sillitoe, B. Smithers, S. Squizzato, G. Sutton, N. Thanki, P. D. Thomas, S. C. E. Tosatto, C. H. Wu, I. Xenarios, L.-S. Yeh, S.-Y. Young and A. L. Mitchell, Nucleic Acids Res., 2017, 45, D190–D199 CrossRef CAS.
- B. Tudzynski, M. Mihlan, M. C. Rojas, P. Linnemannstons, P. Gaskin and P. Hedden, J. Biol. Chem., 2003, 278, 28635–28643 CrossRef CAS.
- M. D. Lebar, J. W. Cary, R. Majumdar, C. H. Carter-Wientjes, B. M. Mack, Q. Wei, V. Uka, S. De Saeger and J. D. Di Mavungu, Fungal Genet. Biol., 2018, 116, 14–23 CrossRef CAS.
- P. Chankhamjon, D. Boettger-Schmidt, K. Scherlach, B. Urbansky, G. Lackner, D. Kalb, H.-M. Dahse, D. Hoffmeister and C. Hertweck, Angew. Chem., Int. Ed., 2014, 126, 13627–13631 CrossRef.
- K. E. Lebe and R. J. Cox, Chem. Sci., 2019, 10, 1227–1231 RSC.
- M. F. Grau, R. Entwistle, Y.-M. Chiang, M. Ahuja, C. E. Oakley, T. Akashi, C. C. C. Wang, R. B. Todd and B. R. Oakley, ACS Chem. Biol., 2018, 13, 3193–3205 CrossRef CAS.
- G. Yang, M. S. Rose, B. G. Turgeon and O. C. Yoder, Plant Cell, 1996, 8, 2139–2150 CAS.
- E. Geiser, S. K. Przybilla, A. Friedrich, W. Buckel, N. Wierckx, L. M. Blank and M. Bölker, Microb. Biotechnol., 2016, 9, 116–126 CrossRef CAS.
- D. W. Brown, S.-H. Lee, L.-H. Kim, J.-G. Ryu, S. Lee, Y. Seo, Y. H. Kim, M. Busman, S.-H. Yun, R. H. Proctor and T. Lee, Mol. Plant-Microbe Interact., 2015, 28, 319–332 CrossRef CAS.
- L. Blank, J. Green and J. R. Guest, Microbiology, 2002, 148, 133–146 CrossRef CAS.
- L. Eichinger, J. A. Pachebat, G. Glockner, M.-A. Rajandream, R. Sucgang, M. Berriman, J. Song, R. Olsen, K. Szafranski, Q. Xu, B. Tunggal, S. Kummerfeld, M. Madera, B. A. Konfortov, F. Rivero, A. T. Bankier, R. Lehmann, N. Hamlin, R. Davies, P. Gaudet, P. Fey, K. Pilcher, G. Chen, D. Saunders, E. Sodergren, P. Davis, A. Kerhornou, X. Nie, N. Hall, C. Anjard, L. Hemphill, N. Bason, P. Farbrother, B. Desany, E. Just, T. Morio, R. Rost, C. Churcher, J. Cooper, S. Haydock, N. van Driessche, A. Cronin, I. Goodhead, D. Muzny, T. Mourier, A. Pain, M. Lu, D. Harper, R. Lindsay, H. Hauser, K. James, M. Quiles, M. M. Babu, T. Saito, C. Buchrieser, A. Wardroper, M. Felder, M. Thangavelu, D. Johnson, A. Knights, H. Loulseged, K. Mungall, K. Oliver, C. Price, M. A. Quail, H. Urushihara, J. Hernandez, E. Rabbinowitsch, D. Steffen, M. Sanders, J. Ma, Y. Kohara, S. Sharp, M. Simmonds, S. Spiegler, A. Tivey, S. Sugano, B. White, D. Walker, J. Woodward, T. Winckler, Y. Tanaka, G. Shaulsky, M. Schleicher, G. Weinstock, A. Rosenthal, E. C. Cox, R. L. Chisholm, R. Gibbs, W. F. Loomis, M. Platzer, R. R. Kay, J. Williams, P. H. Dear, A. A. Noegel, B. Barrell and A. Kuspa, Nature, 2005, 435, 43–57 CrossRef CAS.
- B. Bonsch, V. Belt, C. Bartel, N. Duensing, M. Koziol, C. M. Lazarus, A. M. Bailey, T. J. Simpson and R. J. Cox, Chem. Commun., 2016, 52, 6777–6780 RSC.
- A. Zajc, M. J. Romão, B. Turk and R. Huber, J. Mol. Biol., 1996, 263, 269–283 CrossRef CAS.
- J. I. Jimenez, J. F. Juarez, J. L. Garcia and E. Diaz, Environ. Microbiol., 2011, 13, 1718–1732 CrossRef CAS.
- F. Chardon and C. Damerval, J. Mol. Evol., 2005, 61, 579–590 CrossRef CAS.
- C. A. Cuomo, W. A. Untereiner, L.-J. Ma, M. Grabherr and B. W. Birren, Genome Announc., 2015, 3, e00021-00015 Search PubMed.
- Y.-C. Huang, Y.-H. Chen, S.-R. Lo, C.-I. Liu, C.-W. Wang and W.-T. Chang, Mol. Microbiol., 2004, 53, 81–91 CrossRef CAS.
- D. K. Holm, L. M. Petersen, A. Klitgaard, P. B. Knudsen, Z. D. Jarczynska, K. F. Nielsen, C. H. Gotfredsen, T. O. Larsen and U. H. Mortensen, Chem. Biol., 2014, 21, 519–529 CrossRef CAS.
- K. Katayama, T. Kobayashi, H. Oikawa, M. Honma and A. Ichihara, Biochim. Biophys. Acta Protein Struct. Mol. Enzymol., 1998, 1384, 387–395 CrossRef CAS.
- W. Kim, C.-M. Park, J.-J. Park, H. O. Akamatsu, T. L. Peever, M. Xian, D. R. Gang, G. Vandemark and W. Chen, Mol. Plant-Microbe Interact., 2015, 28, 482–496 CrossRef CAS.
- M. L. Nielsen, L. Albertsen, G. Lettier, J. B. Nielsen and U. H. Mortensen, Fungal Genet. Biol., 2006, 43, 54–64 CrossRef CAS.
- T. J. Carver, K. M. Rutherford, M. Berriman, M.-A. Rajandream, B. G. Barrell and J. Parkhill, Bioinformatics, 2005, 21, 3422–3423 CrossRef CAS.
- M. S. Santisteban, T. Kalashnikova and M. M. Smith, Cell, 2000, 103, 411–422 CrossRef CAS.
- G. Büchi, K. M. Snader, J. D. White, J. Zanos Gougoutas and S. Singh, J. Am. Chem. Soc., 1970, 92, 6638–6641 CrossRef.
- T. Hosoe, J. B. Gloer, D. T. Wicklow, H. A. Raja and C. A. Shearer, Heterocycles, 2010, 81, 2123–2130 CrossRef CAS.
- M. Isaka, M. Tanticharoen and Y. Thebtaranonth, Tetrahedron Lett., 2000, 41, 1657–1660 CrossRef CAS.
Footnotes |
| † Electronic supplementary information (ESI) available. CCDC 2011262–2011264. Crystallographic data for compounds 16, 17 and 18. GenBank accession numbers; MT724049 (zopfiellin BGC) and MT724050 (scytalidin BGC). For ESI and crystallographic data in CIF or other electronic format see DOI: 10.1039/d0sc04309e |
| ‡ These authors contributed equally. |
| § The numbering systems used for the maleidrides varies greatly in the literature and shows no consistency. We propose a common more systematic system based on the size of the ring (1–9, 1–8, 1–7 as appropriate) beginning at the carbon alpha to the maleic anhydride ring, which gives the lowest numbers to the side chains. The maleic anhydride carbons would be numbered with a prime, appropriate to the ring numbering, hence 3′4′ and 8′9′ for byssochlamic acid, and 1′′, 2′′, etc. for the first side chain, numbering from the ring junction and 1′′′, 2′′′, etc. for the second side chain. |
| This journal is © The Royal Society of Chemistry 2020 |