
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Biotechnological approach for improvement of Crambe species as valuable oilseed plants for industrial purposes
Nadia Pushkarova
 * and
Alla Yemets
* and
Alla Yemets
Institute of Food Biotechnology and Genomics, National Academy of Sciences of Ukraine, Osypovskogo Str., 2a, Kyiv 04123, Ukraine. E-mail: pushkarovano@gmail.com; yemets.alla@nas.gov.ua
First published on 2nd March 2022
Abstract
Boosting technological innovation for a sustainable and circular bioeconomy encompasses the use of renewable materials and development of highly effective biotechnological approaches to improve the quality of oilseed crops and facilitate their industrial deployment. The interest in cultivating Crambe as a potential crop is steadily growing due to its low propensity to crossbreeding with other oilseed crops, valuable seed oil composition and a high yield capacity. The main focus is located on Crambe abyssinica as the most adapted into the agriculture and well-studied Crambe species. At the same time, the Crambe genus is one of the most numerous of the Brassicaceae family featuring several underestimated (orphaned) species with useful traits (abiotic stress tolerance, wide range of practical applications). This review features progress in the biotechnological improvement of well-adapted and wild Crambe species starting with aseptic culture establishment and plant propagation in vitro reinforced with the use of genetic engineering and breeding techniques. The aim of the paper is to highlight and review the existing biotechnological methods of both underestimated and well-adapted Crambe species improvment, including the establishment of aseptic culture, in vitro cultivation, plant regeneration and genetic transformation to modify seed oil content and morphological traits of valuable species.
 Nadia Pushkarova | Nadia Pushkarova – a researcher at the Department of Cell Biology and Biotechnology at the Institute of Food Biotechnology and Genomics of the NAS of Ukraine. She received her PhD in Biotechnology in 2017. Her research work focuses on plant biotechnology and the study of underestimated plant species. |
 Alla Yemets | Alla Yemets – Prof., Dr, Head of Cell Biology and Biotechnology Department of the Institute of Food Biotechnology and Genomics of NAS of Ukraine. She is an expert in cell biology and plant biotechnology. Her work is devoted to cellular and molecular regulatory mechanisms of plant development, the influence of abiotic factors on cells, and the role of cytoskeletal components in these processes. Her research includes development and application of genetic engineering methods to obtain genetically modified plants, development of new plant genetic transformation methods, nanobiotechnological approaches for cell biology and biomedicine, and finding new bioactive compounds useful in agriculture and biomedicine. |
1. Introduction
Based on production and circulation of energy, the sustainable bioeconomy is strengthening its position – the biotechnology industry's economy was worth $62,5 billion in 2019 compared to $44,47 billion in 2017.1,2 New policies are being elaborated to reduce carbon emissions, improve resource efficiency, explore renewable energies and develop sustainable agriculture.3,4 The large-scale application of a bioeconomy envisages the use of renewable materials and highly effective biotechnologies to improve the quality of oilseed crops and their industrial implementation.1 Oilseed plants can be used for food or industrial purposes depending on the oil composition and it is essential to prevent the overlapping of crops to maintain high seed oil value. This course is highlighted by the European Commission in European Climate Law and aims to reduce biofuel production from food or feed crops to none by the 2030.4The interest in oilseed crops has grown significantly over the last decade driven not only by spreading the application of the concept of sustainable bioeconomy but also thanks to the growing overall demand due to increasing energy consumption and limited petroleum reserves.5 The industry is primarily interested in the fatty acid composition of the seed oil, specifically in those with the chain length between 12 and 22 carbon atoms: palmitic (16![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :0), stearic (18:0), oleic (18:1 Δ9), linoleic (18:2 Δ9,12), α-linolenic (18:3 Δ9,12,15), lauric (12:0) and erucic (22:1 Δ13) acids. High content of the latter is of particular interest for biofuel production.6,7
:0), stearic (18:0), oleic (18:1 Δ9), linoleic (18:2 Δ9,12), α-linolenic (18:3 Δ9,12,15), lauric (12:0) and erucic (22:1 Δ13) acids. High content of the latter is of particular interest for biofuel production.6,7
Global challenges, such as climate change, deteriorating water and soil conditions, and global population growth are raising a challenge to improve the efficiency of food production. A promising approach to solving this problem is the use of wild, underestimated, or neglected (orphaned) crops in agriculture due to their high nutritional value, high adaptability, and resistance to stress.8–10 Wild Crambe plants are considered as one of the underutilized vegetables and an alternative crop to enhance productivity of agriculture in the abiotic stress regions.11,12 Therefore, it is important to estimate the possibility of wild Crambe species application in agriculture for food, feed and oil production either by cultivation or as a source of valuable traits for improving crops. Although there are several articles highlighting the agronomic and agricultural practices of Crambe,14,15 there are no generalized data on the use of different Crambe species, including wild species, to create in vitro tissue cultures, micropropagation and plant genetic modifications that are very important for further biotechnological improvement of valuable oilseed plants. The review is summarizing the existing biotechnological approaches for improvement of both underestimated and well-adapted Crambe species, and reveals the prospects for their further use in crop biotechnology.
2. Crambe plants: agronomic features and seed oil content
The Crambe genus is the most diverse in the Brassicaceae family, and consists of 44 known species.14,16 Crambe species are widely spread and can grow both at the sea level and at 3800 m above the sea level in Himalayas. They can be found in marine or semi-arid and even dry climatic conditions. According to the geographical distribution this genus can be divided into three groups:17,18•Dendrocrambe DC. – endemic species of the Macaronesian archipelago, northern part of Central Europe;
•Leptocrambe DC. – species of the Mediterranean and Africa;
•Sarcocrambe DC. – species grown in Eurasia up to the Western Himalayas.
Crambe plants are annual and perennial grasses or subshrubs. The stem is glabrous or sparsely pubescent with simple hairs. Basal leaves are large, notched-toothed, pinnately dissected or deeply dissected, usually glabrous or sparsely pubescent, fleshy, vesicular-wrinkled and wavy at the edges on strong long petioles. Stem leaves are much smaller than the basal leaves, softer, usually toothed, petiolate and the upper leaves are small filamentous or absent. The flowers are very numerous14,16 and small, white or yellowish-green in color (Fig. 1). The fruit is an unopened pod. Seeds are 1–3 mm in diameter, brown, greyish-brown or black. Flowering occurs in June.14,18
 | ||
| Fig. 1 Crambe koktebelica (a) and Crambe maritima (b) plants (photo made by Kalista Maria in Karadag mountain range, Crimea, Ukraine). Bar 20 cm. | ||
Crambe basic set of chromosomes is x = 15, plants are always polyploid.19,20 The diploid set of chromosomes (2n) varies from 30 to 150 similarly to polyploid rows of 30, 45, 60, 90, 120 and 150 chromosomes.
Crambe plants are well-adapted in Europe, the USA, China, Brazil and other countries. They are mainly annual plants with short life cycle of 90 days and uniform maturation, which allows mechanical harvesting. Crambe plants are known to tolerate soil salinity, drought and temperature changes.7,21,22 Although there is only one species that is being cultivated at an industrial scale – C. abyssinica – there are also a number of native species that can be a valuable source of the genes required for crop improvement.
Crambe is characterized by a high yield capacity (similar to that of spring rapeseed) and relatively high oil content in the seeds (40%).21 The plants naturally contain up to 60% (C. abyssinica) of erucic fatty acid (22:1 Δ13) which disqualifies them for food production.23 Unlike other widely used oilseed crops (e.g. rapeseed and sunflower), Crambe is at a low risk of outcrossing to its wild relatives, its seed morphology is distinct from other oilseed crops. These traits make it possible to grow oilseed crops for food consumption in the immediate vicinity of Crambe plants without the risk of harming the quality of both food and nonfood crops and outcrossing of different traits among varieties or species.24,25 The main source of erucic acid is high erucic acid rapeseed (HEAR).26 At the same time, rapeseed (canola) is cultivated for food purpose and due to seed and plants similarity the problem of mixing and cross pollination between food and nonfood rapeseed is highly possible.26,27 As erucic acid is harmful to human health when consumed with food and it should not enter the food chain, the Crambe species could be an alternative source of erucic acid that eliminate the problem of outcrossing with rapeseed.13,26
Crambe plants (mainly C. abyssinica) are currently cultivated at an industrial scale in USA, Canada and Europe as they are well suited for oil production.6,27–29 The oil from seeds is used for production of plastic films, adhesives, nylon, thermal insulation, corrosion inhibitors, synthetic rubber and industrial lubricant.30–33 It can also be used for biodiesel production34 due to its high resistance to degradation and oxidation.35 Refined Crambe oil can be used for cosmetics and waxes production.7,24 The waste after seed oil extraction can potentially be used for animal feed as a protein supplement due to the high content of crude protein (up to 45%).31,36–45
3. The use of underestimated wild Crambe species
As mentioned above, despite the fact that Crambe genus is known to possess valuable qualities for agriculture and industry, only one species of the genus, Crambe abyssinica, has been widely adapted in agriculture and cultivated mainly for its oil that is rich in erucic acid.7,46 Crambe from Leptocrambe, Crambe, groups are the most numerous and all the species in Leptocrambe group are shown to have erucic acid at levels comparable to those of C. abyssinica.47There are numerous Crambe species beside C. abyssinica that have been used for food, feed or other purposes.48–51 Out of a wide range of the species the most common ones used for food are C. maritima (Sea Kale), C. cordifolia (Giant Colewort), C. orientalis, C. tataria (Tartar Bread Plant), C. kotschyana,52 C. aspera, C. koktebelica, C. pinnatifida and C. steveniana.51 Crambe leaves and roots are known to have been consumed since ancient times. C. maritima (or Sea Kale) leaves were considered a delicacy in ancient Rome and they were also in France and England in the 19th century.53 In the 1990s, there were attempts to introduce this plant as a vegetable on a large scale in France. Nowadays, Sea Kale is considered a sports food and a food supplement with high crude fiber and protein content.50 Green mass also was shown to have potent antioxidant activity and high polyphenol compounds content depending on the plant development stage54 and a possible application in medicine.51 Also, the antimicrobial activity was shown for underground parts of wild Crambe plants.54
Crambe plants are great for ruminant animal and fish feed as seed meal contains 45–58% of protein with well-balanced amino acid content (especially high levels of lysine and methionine).36–45 It could be used as high value feedstock protein, and the aerial part is excellent for hay due to a high biomass yield containing up to 50% of crude protein.13,55,56
Despite application in food and feed, green mass as well as seed meal of some Crambe species contain glucosinolates that are considered toxic for animal and human health.57 However, physical treatment (freezing or boiling) leads to near complete decomposition of glucosinolates thus making the consumption of sprouts and leaves safe.58 Therefore, reducing levels of glucosinolates or disposing of them completely could make Crambe seed meal suitable for ruminant animals. A potential strategy for this may include reduction of the expression levels of the key genes in the glucosinolates biosynthesis pathway.
4. Biotechnological approach for Crambe improvement
4.1. Crambe in vitro aseptic culture establishment
According to the available literature the establishment of Crambe aseptic culture is carried out mainly through seeds that are characterized with high germination capacity. Seed germination correlates with the seed development phase and environmental conditions.59Despite high germination capacity seed dormancy was also reported for Crambe species.59,60 As recommended by Gutormson et al.,61 application of 0.2% potassium nitrate (KNO3) solution is a possible way for breaking seed dormancy of freshly harvested seeds (it should not be applied for seeds that have been stored for more than nine months).62 The other way of breaking seed dormancy is removal of pericarp which was reported by Nunes et al.62 as a minor limiting factor for Crambe seeds germination. However, pericarp and seed coat removal of C. giberosa was an efficient way to establish high seed germination and greatly accelerate this process in vitro.63 Our previously obtained results support the positive effect of pericarp and seed coat removal for successful and fast in vitro germination of several Crambe species native for Ukraine (C. koktebelica, C. tataria, C. aspera, C. steveniana. C. maritima, C. grandiflora, C.cordifolia, C. mitridatis).64–68 It is necessary to mention that, although pericarp removal can be easily performed at an industrial scale, it could increase the risk of seed damage and spreading of fungal infection, so it should be performed when the risk is justified.
The first step in obtaining an aseptic culture is explant surface sterilization of the plant. The result can vary depending on the explant type and sterilizing agent of choice. Seed surface sterilization is a way to obtain aseptic plant culture in the least damaging way for the plant material. The highest number of aseptic plantlets was obtained from the seeds surface sterilization with diocidum (the exposure time 2–3 minutes)64–68 or commercial bleach solution (exposure time 10–20 minutes).69–71 Also, 3% hydrogen peroxide for 10 minutes was successfully used for that purpose.63 The use of antibiotics for elimination of bacterial contamination was shown to be useful. Immersion in a solution containing penicillin and rifampicin (10 mg L−1) before the application of seed surface sterilization protocol was successful to obtain an aseptic culture.71 After the seed surface sterilization seeds were transferred to culture medium of choice (mainly MS or half strength MS) in culture chamber at 22–25 °C with 16-hour photoperiod.64–71 Time of Crambe aseptic seed germination after the seed surface sterilization procedure varied and lasted from 3 days to a month depending on the species, seed storage time and conditions, and on the seed coat removal.64–73 For aseptic culture establishment explants from aseptic seedlings were taken and transferred to a medium for further cultivation and micropropagation.
Considerable efforts have been made to establish Crambe green shoots surface sterilization. For that purpose, C. gibberosa shoots were soaked in soap solution with Tween80 for 10–15 minutes. Then, the shoots were washed in distilled water, soaked in 0.1% HgCl2 solution for 8 minutes and cut into small nodal explants. Cut ends were paraffined to prevent direct impact of sterilization solution.63 After the surface sterilization, the nodal explants were transferred to MS medium74 containing 6-benzylaminopurine (BA) 2.5 mg L−1 in culture chamber at 23–25 °C for 16 hour long photoperiod. After 6–7 days of the cultivation, the nodal explants turned brown and no regeneration was observed.63
4.2. Crambe micropropagation
Further aseptic plantlets micropropagation is performed mainly on the MS solid medium although the choice of nitrogen and carbon source can influence regeneration of Crambe in vitro. Nitrogen is essential for plants growth and development. Aseptic plantlets uptake nitrogen mainly in the form of NO3− and NH4+ ions therefore its sufficient availability in both forms is important for cell growth and differentiation and the balance of these two nitrogen compounds is needed. Lepoivre,75 MS64–71 or N6 (ref. 75) media have been reported as the most efficient nutrient solutions with optimal mineral composition for Crambe regeneration.75The choice of gelling agents for the medium is also important for successful regeneration in vitro. Gelling of the regeneration medium with agar leads to callogenesis and direct somatic embryogenesis and the use of phytobland contributes to indirect shoots regeneration.73 For higher Crambe regeneration in vitro AgNO3 is also used. It has been found that addition of 5 mg L−1 AgNO3 into the medium improves plantlet regeneration frequency.73,75
Depending on the type of explant and growth regulators content in the medium the plant regeneration frequency varies for each species therefore genetic variations of the plants should be taken into account. Efficient in vitro plant regeneration protocols have been obtained only for several Crambe species – C. abyssinica,73 C. giberosa,63 C. orientalis,70 C. tataria,76 C. maritima.77 Propagation of Crambe plants in vitro was mainly carried out according to the protocols – by direct shoot regeneration of differentiated tissues from different explants:63,70,75–77
•Cotyledons (C. giberosa and C. abyssinica, C. orientalis);
•Hypocotyls (C. abyssinica, C. orientalis);
•Lateral buds (C. giberosa, C. abyssinica);
•Apical meristems (C. abyssinica);
•Leaf parts (C. giberosa, C. tataria, C. abyssinica);
•Root parts (C. maritima, C. tataria, C. giberosa);
•Stem parts (C. abyssinica).
Morphogenic potential of leaf explants was estimated for C. giberosa, C. tataria and C. abyssinica but only the ability for callus formation was observed.63,73,76 For C. abyssinica petiole explants indirect somatic embryogenesis frequency was up to 2.5% while for stem explants it was up to 30%.73
Organogenesis from C. giberosa, C. maritima and C. tataria root explants has been studied.63,76,77 Growth regulators were found to stimulate the morphogenic potential. Growing root explants on MS medium supplemented with a combination of cytokinins and auxins contributed to somatic embryogenesis for C. giberosa (0.5 mg L−1 of BA),63 for C. maritima (2.0 mg L−1 of IAA and 0.8 mg L−1 of kinetin),77 and for C. tataria (NAA 1–2 mg L−1 and BA 1–2 mg L−1).76
The use of root explants for plant propagation of C. giberosa, C. maritima and C. tataria in aseptic culture proved to be quite effective provided that the appropriate amount of NAA and BA was added to the medium.
Intensive callogenesis was also shown for C. tataria leaf explants. It was noted76 that both cytokinins and auxins were essential for callus tissue formation (1 mg L−1 2,4-D induced callogenesis on leaf explants), while NAA was less effective than 2,4-D, and BAP, in turn, was less effective than kinetin. Thus, the highest intensity of callus tissue growth on leaf explants was observed on medium with 2 mg L−1 of kinetin and 1 mg L−1 of 2,4-D. C. tataria also showed a low callogenesis activity of root explants, compared to leaf explants. For root explants, the highest activity of callogenesis was observed with the use of medium supplemented with 2 mg L−1 of 2,4-D.76
4.3. In vitro plant regeneration of wild Crambe species
Morphogenic potential of different types of explants of several Crambe species was previously tested by us (Table 1). This research was performed to estimate the possible use of root, leaf and petiole explants for in vitro propagation of Crambe plants that are endangered in Ukraine, as well as in other counties, and to obtain the basis for Crambe gene pool improvement.64–68,78| Species | Lateral bud explants | Leaf explants | Petiole explants | Root explants | References |
|---|---|---|---|---|---|
| C. koktebelica | BA 1 mg L−1 | BA 5 mg L−1 + NAA 0.5 mg L−1 | BA 2.5 mg L−1 + NAA 0.1 mg L−1 | BA 1 mg L−1 + NAA 0.1 mg L−1 | 64, 67, and 78 |
| C. tataria | BA 0.3 mg L−1 | BA 1 mg L−1 + NAA 0.1 mg L−1 | BA 1 mg L−1 + NAA 0.1 mg L−1 | Kinetin 1 mg L−1 + NAA 0.5 mg L−1 | 65, 76, and 78 |
| C. aspera | Kinetin 0.5 mg L−1 | BA 5 mg L−1 + NAA 0.5 mg L−1 | BA 5 mg L−1 + NAA 1 mg L−1 | BA 1 mg L−1 + NAA 0.1 mg L−1 | 68 and 78 |
| C. steveniana | BA 0.6 mg L−1 | BA 1 mg L−1 + NAA 0.1 mg L−1 | BA 2.5 mg L−1 + NAA 0.1 mg L−1 | Kinetin 1 mg L−1 + NAA 0.1 mg L−1 | 78 |
| C. maritima | BA 1 mg L−1 | BA 2.5 mg L−1 + NAA 0.5 mg L−1 | BA 2.5 mg L−1 + NAA 0.1 mg L−1 | BA 1 mg L−1 + NAA 0.1 mg L−1 | 78 |
Morphogenic potential of root explants of wild Crambe species has been researched and showed rather low regeneration frequency for four species (Fig. 2): C. koktebelica – 20% (BA 1 mg L−1 and NAA 0.1 mg L−1), C. aspera – 10% (BA 1 mg L−1 and NAA 0.1 mg L−1), C. tataria – 30% (Kinetin 1 mg L−1 and NAA 0.5 mg L−1) and C. steveniana – 40% (Kinetin 1 mg L−1 and NAA 0.1 mg L−1) (Table 1). The highest micropropagation rates were observed in the case of cultivation with low cytokinin and auxin content in the medium. Plant regeneration from C. koktebelica and C. aspera roots took place only on the medium with BA. C. maritima root explants had the highest micropropagation rates on the medium with either BA or kinetin, all of its explants formed plantlets on the MS medium with 1 mg L−1 of BA and 0.1 mg L−1 of NAA.
 | ||
| Fig. 2 Plantlets regeneration on petiole explants: (a) C. koktebelica; (b) C. tataria; (c) C. aspera; (d) C. steveniana; (e) C. maritima. Bar 1 cm. | ||
The conducted experiments have demonstrated that regeneration frequency of leaf explants was higher than that of root explants. New plantlets for C. koktebelica, C. tataria, C. aspera, C. steveniana and C. maritima were formed on the medium containing either BA and NAA, or kinetin and NAA, however, the former combination (BA and NAA) showed the highest propagation rates. The relation between the regeneration frequency and auxin content in the medium was also established, lower NAA concentration (0.1–0.5 mg L−1) made for higher propagation rates. Thus, the highest regeneration frequency from leaf explants was as follows: 28% for C. koktebelica (BA 5 mg L−1 and NAA 0.5 mg L−1), 38% for C. tataria (BA 1 mg L−1 and NAA 0.1 mg L−1), 50% for C. aspera (BA 5 mg L−1 and NAA 0.5 mg L−1), 80% for C. steveniana (BA 1 mg L−1 and NAA 0.1 mg L−1) and 100% for C. maritima (BA 2.5 mg L−1 and NAA 0.5 mg L−1) (Table 1).
We have also established that NAA concentration in the medium was an important regulatory factor for both petiole and leaf explants regeneration rate, the concentration 0.1–0.5 mg L−1 of NAA resulted in the higher propagation rates. Plantlets from petioles were formed on the medium with either BA or kinetin (combined with NAA). 100% regeneration frequency occurred for C. tataria (BA 1 mg L−1 and 0.1 mg L−1), C. aspera (BA 5 mg L−1 and 1 mg L−1), C. steveniana (BA 2.5 mg L−1 and NAA 0.1 mg L−1) and for C. maritima (BA 2.5 mg L−1 and NAA 0.1 mg L−1). The highest propagation rates for C. koktebelica petiole explants (60%) were noted on the medium with 2.5–5 mg L−1 of BA and 0.1 mg L−1 of NAA (Table 1).64–68,78
Our previously conducted research provides a full way from a Crambe seed to numerous plants in greenhouse via in vitro propagation with rooting and acclimatization of plants for several species (Fig. 3). It can therefore be inferred, that a platform for further biotechnological improvement of Crambe species is established and is applicable to other relevant methods. Underestimated Crambe species are often endemics and endangered14,79,80 therefore the problem with reproduction in their natural habitats or law restrictions due to the threatened status of plants is present. In vitro propagation could provide with rapid multiplication of plants that have characteristics of mother plant without considerable damage to the population in their habitats. Crambe genetic breading for increasing oil and other compounds production requires high efficiency of biotechnological steps.75 Establishment of propagation protocols for each of Crambe species is fairly important. Propagation protocols can be further used as a platform to transfer desirable traits (Fig. 4) via cellular and genetic engineering approaches or to generate new lines with desirable traits due to somaclonal variability that can occur by chance when plant tissues are cultured in vitro.81 By combining existing propagation protocols for underestimated species with the knowledge on genetic constructs used for C. abyssinica for enhancing desirable traits the new varieties could be obtained.
 | ||
| Fig. 3 C. aspera in vitro micropropagation and plant adaptation to greenhouse conditions. | ||
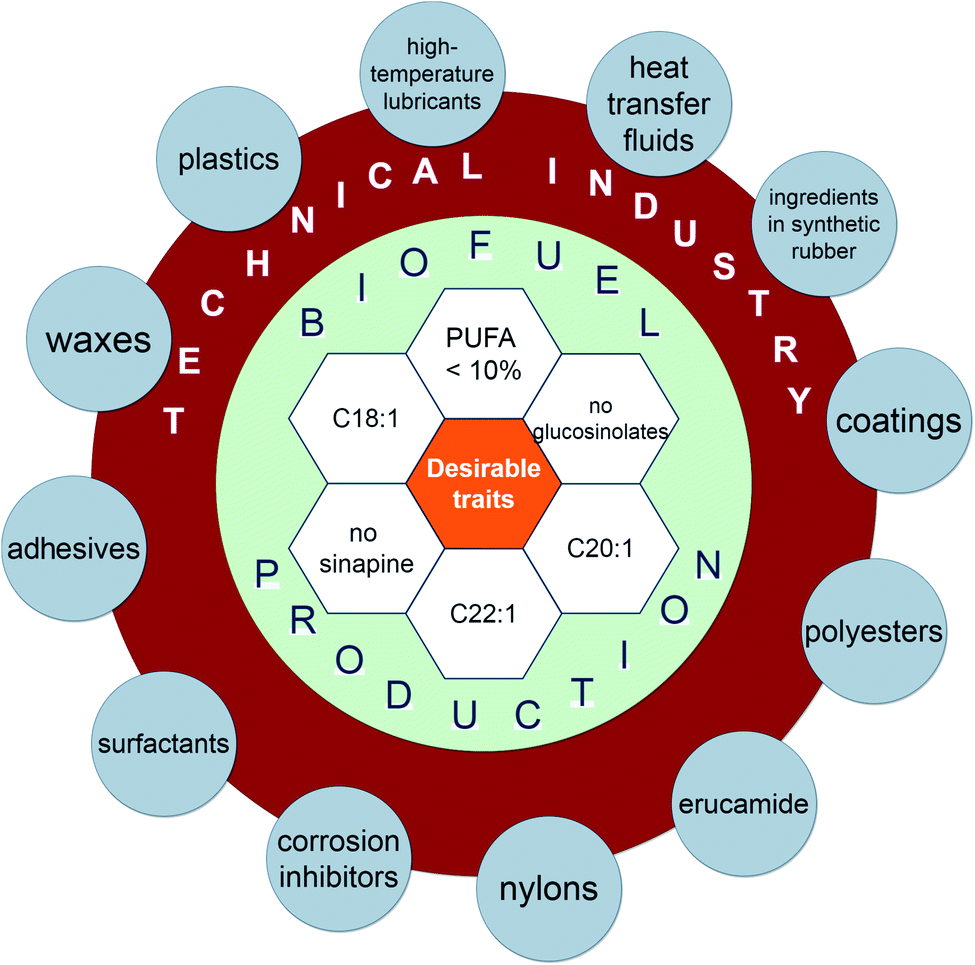 | ||
| Fig. 4 The various components of Crambe species for possible applications in biofuel production and technical industry. | ||
4.4. Somatic hybridization as a method for Crambe improvement
Somatic hybridization is a method of cellular genome manipulation by protoplasts fusion of two different species to form a new hybrid plant with combined features. It may be intraspecific, interspecific, intrageneric and intergeneric. Somatic hybridization can be performed between different varieties of one species and between different species even not closely related. As a result, the hybrids similar to polyploid plants can be obtained with the chromosomes of both plants. Plant protoplasts fusion as a method of somatic hybridization is an essential approach to overcome sexual incompatibility between different plant species thus providing a tool for new genetic traits formation. The most common recipients for Crambe genome transfer via somatic hybridization are Brassica spp., namely B. napus, B. campestris, B. juncea.82,83 Major limitations were found in the intergeneric cross process between Brassica spp. and C. abyssinica such as prefertilization incompatibility and embryo abortion at multi-cellular globular stage.82 To overcome an embryo abortion ovary culture was successfully used and B. juncea × C. abyssinica hybrids were obtained.80 Wang et al.82 demonstrated a successful transfer of new allelic variants of the FAE1 (fatty acid elongation gene controlling erucic acid biosynthesis) from C. abyssinica into B. napus via somatic hybridization. Overexpression of the C. abyssinica FAE1 gene in B. carinata resulted in a substantial increase in the content of erucic acid in seeds compared to the wild type control.84UV-irradiated (0.10 J cm−3 dose) protoplasts from C. abyssinica leaves were mixed in vitro with B. napus protoplasts (in 1.2:1 ratio) and treated with 40% polyethylene glycol, then cultivated in the dark for further microcalli formation and hybrid plant regeneration. Obtained asymmetric somatic hybrids had from 2 to 40 more chromosomes than expected for B. napus (38 chromosomes) and showed the presence of some characteristic bands from C. abyssinica (confirmed by the amplified fragment-length polymorphism analysis (AFLP)). The obtained hybrids had high pollen viability, could be fertilized and set seeds. The subsequent seed oil content estimation showed several asymmetric somatic hybrids with an increased erucic acid content and seed set.17
Intraspecific hybridization as a way of transferring desirable traits of related wild species (diseases resistance, abiotic stress resistance, higher yield etc.) to well-adapted and cultured crop C. abyssinica85–88 was used by Du et al.89 Though, this approach is limited due to the sexual incompatibility between species that leads to low fertility of hybrids or the abortion of embryo at early development stages, such obstacles can be overcome by protoplast fusion and embryo rescue.90,91 Hybrids of C. abyssinica, C. hispanica and C. kralikii can be obtained with or without embryo rescue.89
4.5. Agrobacterium-mediated genetic transformation of Crambe
Agrobacterium-mediated transformation is a widely used method of plants genetic engineering due to its high efficiency, which is influenced by several factors such as plant donor health, stage of the donor material, vector type, regeneration and selection conditions efficiency.23,72,92,92 As Crambe plants are considered highly potential oilseeds and a source of desirable genes for crop improvement, its genetic transformation is aimed to change the seed fatty acid composition and increase the very long chain fatty acids (VLCFA) content in seeds.84,94–96Genetic modification of Crambe plants became possible after the development of the first successful plant regeneration and the transformation protocol mediated by Agrobacterium.93 To overcome the so-called bottleneck in erucic acid accumulation97 in attempt to decrease polyunsaturated fatty acids content in Crambe seeds, up to four gene-combined constructs were created and used for transformation.96 The first attempt to apply gene stacking strategy for increasing erucic acid content in Crambe was performed by Li et al.23,72 For this purpose, they used vectors pHAN, pWatergate and three-gene construct harbouring the LdLPAAT (lysophosphatidate acyltransferase gene from Limnanthes douglasii),98 CaFAD2-RNAi (fatty acid desaturase 2 gene) and BnFAE1 (fatty acid elongase 1 gene from B. napus) genes driven under the napin promoter and with nptII selectable marker gene23 and binary vector pCAMBIA carrying BnFAE1 and LdLPAAT with hpt selectable marker gene.72 It has been shown that incorporation of FAD2-RNAi together with the previously mentioned genes resulted in the increase of erucic acid amount in Crambe seed oil up to 70%.23,72,94
Crambe gene could be a useful source for improving oilseed crops by cross-species silencing.99 RNAi-silencing construct containing Crambe FAD2, FAD3 and FAE1 genes was used for silencing the genes of related Arabidopsis thaliana resulting in fatty acid content alteration. Expression of CaFAD2-FAE1 gene silencing constructs decreased cis-11 eicosenoic (20:1) and linoleic (18:2 Δ9,12) and CaFAD3-FAE1 decreased α-linolenic (18:3 Δ9,12,15) fatty acid content in seed oil.99 The same constructs were used for developing ultrahigh oleic oil content into C. abyssinica resulting in a significant increase in oleic acid (18:1 Δ9) content wherein, and this trait was stable during several generations.96 Suppressing of the lysophosphatidic acid acyltransferase LPAAT2 expression was also performed via RNAi targeting with CaFAD2 and CaLPAAT2 genes from Crambe.26
Crambe seed oil contains almost 70% of VLCFAs which make it an attractive source for the biotechnological production of industrial oils by overlaying the wax ester biosynthetic pathway from jojoba onto the existing triacylglycerol biosynthetic pathways of C. abyssinica. Wax ester biosynthesis requires the fatty acid esterification from an acyl-CoA substrate to a fatty alcohol, bypassing the fatty acid incorporation onto glycerol backbones to form triacylglycerols. For this purpose, ScFAR and ScWS cDNAs were co-expressed under control of strong seed-specific promoters in Crambe resulting in successful tailoring wax ester profiles.24 Despite promising results in altering FA content for wax esters, a production field and greenhouse trials showed that transgenic lines expressing ScWS and ScFAR genes provided normal growth of transgenic plants but with a slightly reduced seed yield, oil content and germination rate compared to the wild type with delayed flowering and fruit set. Crambe lines selected for a field testing had approximately 25% of the oil as wax esters, with the remainder in the form of TAG.24,100
4.6. Crambe hairy roots culture establishment
Genetic transformation by means of Agrobacterium rhizogenes results in hairy roots culture formation that is caused by root loci genes incorporation into the plant DNA. Hairy root culture is characterized by rapid hormone-independent growth, lateral branching, high genetic and biochemical stability and can produce valuable secondary metabolites non inherent for the initial plant.101,102 This technology is used for secondary metabolites production, plants biochemical properties study and could be used at an industrial scale in bioreactors.103 The possibility to use hairy root culture for oil production in bioreactors was also reported.104,105Crambe hairy root culture was established using leaf and cotyledons explants inoculated with A. rhizogenes A4 and 15834 strains by two methods, needle inoculation and sonication.106 It should be noted that addition of 200 mM acetosyringone (4-acetyl-2,6-dimethoxyphenol) to the growth medium, used for A. rhizogenes and inoculated plants explants growth, increased transformation efficiency. Hairy root culture that was incubated for more than three weeks showed symptoms of aging (lack of growth and culture browning due to intensive phenolic compounds excretion) but admixing polyvinylpyrrolidone reduced the culture aging and intensified its development.106 The hairy root culture establishment was studied with the use of cotyledons and leaf explants and it was shown that the leaf explants were of better choice for Crambe transformation resulting in 16% transformation efficiency.106 This work106 was the first attempt to obtain stable C. abyssinica hairy roots culture that could be characterized by genetic stability, rapid growth on hormone-free medium and possible high polyunsaturated fatty acids production. These traits make it possible to use hairy roots culture in bioreactors for oil production. For this purpose, hairy roots fatty acids content of different Crambe species should be studied further.
Fatty acids content in the hairy root culture was similar to that of non-transformed roots but differed only in terms of quantity. Dominating fatty acids of A. rhizogenes-induced roots were α-linolenic (18:3 Δ9,12,15), palmitic (16:0), and linoleic (18:2 Δ9,12) acids. Diacylglycerols, free fatty acids, triacylglycerols, and sterol esters, found in root culture and polar lipids, were the dominant class of lipids.105 The obtained results provide the protocol of hairy root culture establishment and reveal some features of anabolism and catabolism of Crambe root lipids.
Further attempts of improving Crambe hairy roots lipid metabolism were focused on introducing genes coding for the fatty acyl-CoA reductases from A. thaliana (AtFAR5 gene) and Simmondsia chinensis (ScFAR gene). A. rhizogenes A4 and ATCC 15834 stains carrying binary plasmids pBIN-AtFAR5, pGW-AtFAR5, pBIN-ScFAR and pGW-ScFAR were applied for the transformation of C. abyssinica leaf explants.107 The reported transformation efficiency was 35–45% and the obtained hairy roots assessment showed different fatty alcohol profiles – culture containing AtFAR5 gene was unable to produce fatty alcohols but for the lines carrying ScFAR gene stearyl alcohol (18:0-OH), arachidyl alcohol (20:0-OH) and behenyl alcohol (22:0-OH) production was noted.107
Hairy root culture is a valuable source for fatty acids production but further research should be conducted to estimate the full potential of this system and to see if it could compete with the conventional seed oil production.
4.7. The potential of CRISPR/Cas9 gene editing for improvement of Crambe seed oil content and other traits
Due to a continuously growing demand for food and energy new ways to increase production of vegetable renewable oil are required. Genome editing techniques, such as clustered regularly interspaced short palindromic repeats-associated protein (CRISPR/Cas9), have emerged as a powerful, highly specific and eco-friendly tool for crops improvement.108,109 This system comprises Cas nuclease which makes double strand DNA breaks and a small non-coding single-guide RNA that leads Cas to the destinated genomic locus (DBS) are then repaired by plant inherent cell repair mechanisms.110Crambe plants are a dedicated source of fatty acids that are accumulated in the form of triacylglycerols in seeds.111 Triacylglycerol formation is a complex process that could be altered at certain steps to overcome the bottleneck in erucic acid accumulation97 and change polyunsaturated fatty acid content in Crambe seed oil. Considerable efforts have been made for oilseed crops gene editing by means of CRISPR/Cas9 with targeting genes that are involved in triacylglycerol synthesis: lysophosphatidic acid acyltransferase,112 fatty acid desaturase,113–115 fatty acid elongase,116 diacylglycerol acyltransferase.117 At the same time, there is no data available on Crambe gene editing via CRISPR/Cas9 system, therefore the study of this system application to Crambe is of particular interest.
5. Conclusions
Crambe plants have great potential and a wide range of technological application. The considerable efforts have been made to produce biotechnological systems for Crambe improvement starting from in vitro regeneration protocols and somatic hybridization to Agrobacterium-mediated genetic transformation with genes regulating lipid production. Further research in this field could provide a useful genetic material for crop improvement as well as new systems for producing valuable fatty acids.Author contributions
Nadia Pushkarova carried out the literature search, summarized data and wrote the paper. Alla Yemets reviewed and edited the manuscript, analyzed the manuscript contents and made the manuscript corrections. All authors have read and approved the final manuscript version.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This work was supported by the Grant of National Academy of Sciences of Ukraine for Young Scientists “Physiological and cell mechanisms of salt tolerance of wild Crambe species” [No. 0121U112000, 2021–2022]. The authors are grateful to Dr V. Kyrylenko (Institute of Food Biotechnology and Genomics, National Academy of Sciences of Ukraine, Kyiv) for final improving of English version of the manuscript.Notes and references
- N. Scarlat, J.-F. Dallemand, F. Monforti-Ferrario and V. Nita, Environ. Dev., 2015, 15, 3–34, DOI:10.1016/j.envdev.2015.03.006.
- Indian Bioeconomy Report 2021, https://birac.nic.in/webcontent/1579089535_India_BioEconomy_Report_2019.pdf, accessed December 2021 Search PubMed.
- European Commission, Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions, 2014, https://www.eea.europa.eu/policy-documents/communication-from-the-commission-to-1, accessed December 2021 Search PubMed.
- European Commission, Directive (EU) 2018/2001 of the European Parliament and of the Council of 11 December 2018 on the promotion of the use of energy from renewable sources, 2018, https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32018L2001%26from=fr, accessed December 2021 Search PubMed.
- R. Dolah, R. Karnik and H. Hamdan, Sustainability, 2021, 13, 10210, DOI:10.3390/su131810210.
- A. Carlsson, Biochimie, 2009, 91, 665–670, DOI:10.1016/j.biochi.2009.03.021.
- S. Falasca, N. Flores, M. Lamas, S. Carballo and A. Anschau, Int. J. Hydrogen Energy, 2010, 35, 5808–5812, DOI:10.1016/j.ijhydene.2010.02.095.
- K. Gruber, Nat, 2017, 544, 8–10, DOI:10.1038/544S8a.
- A. Singh, P. Dubey and P. Abhilash, Curr. Sci., 2018, 155, 611–613, DOI:10.18520/CS/V115/I4/611-613.
- S. Padulosi, J. Thompson and P. Rudebjer, Fighting Poverty, Hunger and Malnutrition with Neglected and Underutilized Species (NUS): Needs, Challenges and the Way Forward, Bioversity International, Rome, 2013. DOI:10.13140/RG.2.1.3494.3842.
- A. Hossain, T. Islam, S. Maitra, D. Majumder, S. Garai, M. Mondal, A. Ahmed, A. Roy, M. Skalicky, M. Brestic, and T. Islam, in Neglected and Underutilized Crops – Towards Nutritional Security and Sustainability, ed. S. M. Zargar, A. Masi, and R. K. Salgotra, Springer, Singapore, 2021. DOI:10.1007/978-981-16-3876-3_1.
- A. Singh, P. K. Dubey, R. Chaurasia, R. K. Dubey, K. K. Pandey, G. S. Singh and P. C. Abhilash, Agronomy, 2019, 9, 491, DOI:10.3390/agronomy9090491.
- D. Samarappuli, F. Zanetti, S. Berzuini and M. T. Berti, Agronomy, 2020, 10, 1380, DOI:10.3390/agronomy10091380.
- F. Branca and E. Cartea, in Wild Crop Relatives: Genomic and Breeding Resources, Oilseeds, ed. C. Kole, Springer, Heidelberg, 2011, pp. 17–36 Search PubMed.
- D. Bassegio, M. D. Zanotto, R. F. Santos, I. Werncke, P. P. Dias and M. Olivo, Renewable Sustainable Energy Rev., 2016, 66, 311–321, DOI:10.1016/j.rser.2016.08.010.
- A. O. Prina, An. Inst. Bot. A. J. Cavanilles, 2009, 66, 7–24, DOI:10.3989/ajbm.2186.
- Y. P. Wang, K. Sonntag and E. Rudloff, Theor. Appl. Genet., 2003, 106, 1147–1155, DOI:10.1007/s00122-002-1176-x.
- O. Appel and I. Al-Shebaz, in Flowering plants-Dicotyledons. Malvales Capparales and non-betalain Caryophyllales, ed. K. Kubitzki, Springer, Berlin, 2003, pp. 75–174 Search PubMed.
- K. Marhold and J. Lihova, Plant Syst. Evol., 2006, 259, 143–174, DOI:10.1007/s00606-006-0417-x.
- S. I. Warwick and I. A. Al Shehbaz, Plant Syst. Evol., 2006, 259, 237–248, DOI:10.1007/s00606-006-0421-1.
- M. F. Silva, E. F. Araújo, L. J. Silva, H. T. R. Amaro, L. A. S. Diaz and D. C. F. S. Dias, Cienc. Agrotecnol., 2019, 43, e025418, DOI:10.1590/1413-7054201943025418.
- G. Castleman, S. Paymer and C. Greenwood, in Proceedings of the 10th international rapeseed congress, Canberra, Australia, 1999, pp. 26–29 Search PubMed.
- X. Li, G. van Loo, J. Gruber, J. Fan, R. Guan, M. Frentzen, S. Stymne and L. H. Zhu, Plant Biotechnol. J., 2012, 10, 862–870, DOI:10.1111/j.1467-7652.2012.00709.x.
- L. H. Zhu, in Industrial Oil Crops, ed. T. A. McKeon, D. G. Hayes, D. F. Hildebrand, and R. J. Waselake, Elsevier Inc., Cambridge, 2016, pp. 195–205. DOI:10.1016/B978-1-893997-98-1.00007-5.
- F. Zanetti, D. Scordia, T. Vamerali, V. Copani, C. Dal Cortivo and G. Mosca, Ind. Crops Prod., 2016, 90, 152–160, DOI:10.1016/j.indcrop.2016.06.023.
- W. Qi, I. E. M. Tinnenbroek-Capel, E. M. J. Salentijn, Z. Zhao, B. Huang, J. Cheng, H. Shao, R. G. F. Visser, F. A. Krens and E. N. van Loo, Land Degrad. Dev., 2018, 29, 1096–1106, DOI:10.1002/ldr.2847.
- A. Hebard, in Industrial Oil Crops, ed. T. A. McKeon, D. G. Hayes, D. F. Hildebrand, and R. J. Weselake, AOCS Press, Winston-Salem, NC, 2016, pp. 343–358 Search PubMed.
- M. J. Stolarski, M. Krzyz˙aniak, J. Tworkowski, D. Załuski, J. Kwiatkowski and S. Szczukowski, Ind. Crops Prod., 2019, 137, 386–395, DOI:10.1016/j.indcrop.2019.05.047.
- E. Costa, M. F. Almeida, C. Alvim-Ferraz and J. M. Dias, Ind. Crops Prod., 2019, 129, 51–58, DOI:10.1016/j.indcrop.2018.11.032.
- S. L. Falasca, N. Flores, M. C. Lamas, S. M. Carballo and A. R. Anschau, Int. J. Hydrogen Energy, 2010, 35, 5808–5812, DOI:10.1016/j.ijhydene.2010.02.095.
- K. D. Carlson, J. C. Gardener, V. L. Anderson, and J. J. Hanzel, in Progress in new crops, ed. J. Janick, ASHS Press, Alexandria, 1996, pp. 306–322 Search PubMed.
- W. R. Newson, M. L. Prieto-Linde, R. Kuktaite, M. S. Hedenqvist, M. Gällstedt and E. Johansson, Ind. Crops Prod., 2017, 97, 591–598, DOI:10.1016/j.indcrop.2016.12.037.
- N. Volkova, X. Li and L. H. Zhu, et al., Sustainable Chem. Processes, 2016, 4, 1, DOI:10.1186/s40508-016-0045-x.
- G. Tavares, T. Massa, J. Gonçalves, C. Silva and W. Dos-Santos, Renewable Energy, 2017, 111, 659–665, DOI:10.1016/j.renene.2017.04.065.
- S. Lalas, O. Gortzi, V. Athanasiadis, E. Dourtoglou and V. Dourtoglou, J. Am. Oil Chem. Soc., 2012, 89, 2253–2258, DOI:10.1007/s11746-012-2122-y.
- R. De Goes, R. A. Patussi, J. R. Gandra, A. F. Branco, T. J. D. Cardoso, M. V. M. de Oliveira, R. T. de Oliveira and C. J. D. Souza, J. Biosci., 2017, 33, 113–120, DOI:10.14393/BJ-v33n1a2017-33105.
- R. H. De Goes, M. M. V. Carneiro, M. P. Osmari, K. A. De Souza, R. T. De Oliveira and C. J. D. Souza, Acta Sci., Anim. Sci., 2018, 40, e37171, DOI:10.4025/actascianimsci.v40i1.37171.
- R. H. De Goes, R. Patussi, A. Branco, M. Osmari, J. Gandra, W. Zeviani, L. Bezerra and R. Oliveira, Ital. J. Anim. Sci., 2019, 18, 316–327, DOI:10.1080/1828051X.2018.1528898.
- M. M. Y. Carneiro, R. H. De Goes, L. H. Z. Da Silva, A. R. M. Fernandes, R. T. De Oliveira, C. A. L. Cardoso, A. S. O. Hirata and R. Bras, Zootec, 2016, 45, 319–327, DOI:10.1590/S1806-92902016000600006.
- E. B. Canova, M. S. Bueno, H. L. Moreira, R. Possenti and P. Bras, Cienc. Agrotecnol., 2015, 39, 75–81, DOI:10.1590/S1413-70542015000100009.
- L. C. V. Ítavo, A. D. V. de Souza, S. P. Fávaro, C. C. B. F. Ítavo, H. V. Petit, A. M. Dias, M. G. Morais, R. G. Coelho, F. A. Reis and J. A. A. Costa, et al., Anim. Feed Sci. Technol., 2016, 216, 40–48, DOI:10.1016/j.anifeedsci.2016.02.015.
- M. A. Syperreck, I. Y. Mizubuti, E. S. Pereira, E. L. A. Ribeiro, E. L. T. Peixoto, P. G. Pimentel, A. L. C. Franco, F. L. Massaro, R. M. Brito and A. R. P. Parra, Semina: Cienc. Agrar., 2016, 37, 2633–2640, DOI:10.5433/1679-0359.2016v37n4Supl1p2633.
- N. M. Lovatto, B. B. Loureiro, D. Pianesso, T. J. Adorian, F. R. Goulart, C. S. Speroni, A. B. B. Bender, J. Müller and L. P. Da Silva, An. Acad. Bras. Ciênc., 2018, 90, 3781–3790, DOI:10.1590/0001-3765201820170991.
- A. M. Cardoso, S. A. D. C. Araujo, N. C. Rocha, F. M. Domingues, J. C. de Azevedo and L. D. A. Pantoja, Acta Sci., Anim. Sci., 2016, 38, 375–382, DOI:10.4025/actascianimsci.v38i4.31828.
- J. Issakowicz, M. C. Bueno, C. M. P. Barbosa, E. B. Canova, H. L. Moreira, A. T. Geraldo and A. C. K. Sampaio, Anim. Prod. Sci., 2017, 57, 785–792, DOI:10.1071/AN15360.
- M. Krzyaniak, M. Stolarski, M. Ânieg and E. Alexopoulou, Environ. Biotechnol., 2014, 9, 72–80, DOI:10.14799/ebms220.
- E. Leppik and G. White, Euphytica, 1975, 24, 681–689, DOI:10.1007/BF00132906.
- S. Duhoon and M. Koppar, Genet. Resour. Crop Evol., 1998, 45, 317–323, DOI:10.1023/A:1008646501454.
- N. Ul’chenko, N. Bekker, A. Glushenkova and G. Akhmedzhanov, Chem. Nat. Compd., 2001, 37, 285–286, DOI:10.1007/s10600-011-9770-2.
- Sea Kale and Other Crambes, https://www.cultivariable.com/instructions/other-vegetables/how-to-grow-sea-kale-and-other-crambes/, accessed December 2021 Search PubMed.
- M. Kalista, Agrobiodiv. Impr. Nut. Health Life Qual., 2017, 1, 216–220, DOI:10.15414/agrobiodiversity.2017.2585-8246.216-220.
- E. Rudloff and Y. Wang, in Wild Crop Relatives: Genomic and Breeding Resources, ed. C. Kole, Springer, Berlin, 2011, pp. 97–116 Search PubMed.
- J. M. Renfrew and H. Sanderson, in The cultural history of plants, ed. G. Prance, and M. Nesbitt, Routledge, Renfrew, 2005, pp. 97–131 Search PubMed.
- O. Vergun, D. Rakhmetov, O. Shymanska, O. Bondarchuk, E. Ivanišová, M. Kačaniová, O. Grygorieva and J. Brindza, Acta Sci. Pol., Technol. Aliment., 2021, 20, 197–211, DOI:10.17306/J.AFS.2021.0927.
- V. L. Anderson, W. D. Slanger, S. L. Boyles and P. T. Berg, J. Anim. Sci., 1993, 71, 2608–2613, DOI:10.2527/1993.71102608x.
- A. Steg, V. A. Hindle and Y.-G. Liu, Anim. Feed Sci. Technol., 1994, 50, 87–99, DOI:10.1016/0377-8401(94)90011-6.
- J. W. Fahey, A. T. Zalcmann and P. Talalay, Phytochemistry, 2001, 56, 5–51, DOI:10.1016/s0031-9422(00)00316-2.
- L. Yong-Gang, A. Steg and V. Hindle, Anim. Feed Sci. Technol., 1993, 41, 133–147 CrossRef.
- J. Kwiatkowski, M. Krzyzaniak, D. Zaluski, M. J. Stolarski and J. Tworkowski, Ind. Crops Prod., 2020, 145, 111977, DOI:10.1016/j.indcrop.2019.111977.
- L. M. Costa, O. Resende, D. N. Gonçalves, K. A. Souza, J. F. Sales and J. R. Donadon, Acta Sci., Agron., 2011, 34, 213–218, DOI:10.4025/actasciagron.v34i2.12482.
- T. J. Gutormson, K. L. Larson and T. Waqar, J. Seed Technol., 1992, 6, 17–23, DOI:10.2307/23432785.
- A. S. Nunes, L. C. F. de Souza, S. P. Q. Scalon and J. Pagnoncelli, Semin Cienc Agrar, 2015, 36, 1775–1782, DOI:10.5433/1679-0359.2015v36n3Supl1p1775.
- V. K. Magomedalieva, Fund. Res., 2013, 10, 114–118 Search PubMed.
- N. O. Pushkarova, M. S. Kalista, M. S. Kharhota, D. B. Rakhmetov and M. V. Kuchuk, Biotechnol. Acta, 2016, 9, 19–27, DOI:10.15407/biotech9.04.019.
- N. O. Pushkarova, M. S. Kalista, M. S. Kharhota, D. B. Rakhmetov and M. V. Kuchuk, Potr. S. J. F. Sci., 2016, 10, 494–498, DOI:10.5219/646.
- N. O. Pushkarova, T. M. Kyrpa-Nesmiian and M. V. Kuchuk, Visnik ukrains'kogo tovaristva genetikiv i selekcioneriv, 2019, 17, 172–178, DOI:10.7124/visnyk.utgis.17.2.1218.
- N. O. Pushkarova, O. R. Lakhneko, V. B. Belokurova, B. V. Morgun and M. V. Kuchuk, Cytol. Genet., 2018, 52, 269–275, DOI:10.3103/S0095452718040096.
- N. O. Pushkarova, O. R. Lakhneko, B. V. Morgun, M. V. Kuchuk, Ya. B. Blume and A. I. Yemets, Biopolym. Cell, 2019, 35, 118–128, DOI:10.7124/bc.00099D.
- S. Chhikaraa, I. Duttaa, B. Paulosea, P. K. Jaiwal and O. P. Dhankher, Ind. Crops Prod., 2012, 37, 457–465, DOI:10.1016/j.indcrop.2011.07.021.
- Y. Ceylan, F. A. Özdemir and A. S. Bülbü, J. Anim. Plant. Sci., 2017, 27, 1671–1677 CAS.
- E. Werner, C. Milanez, A. Gontijo, T. Soares and J. Amaral, Afr. J. Biotechnol., 2016, 15, 722–730, DOI:10.5897/AJB2015.14965.
- X. Li, J. Fan, J. Gruber, R. Guan, M. Frentzen and L. H. Zhu, Front. Plant Sci., 2013, 4, 162, DOI:10.3389/fpls.2013.00162.
- W. Qi, I. E. M. Tinnenbroek-Capel, J. G. Schaart, B. Huang, J. Cheng, R. G. F. Visser, E. N. Van Loo and F. A. Krens, BMC Plant Biol., 2014, 14, 235, DOI:10.1186/s12870-014-0235-1.
- T. Murashige and F. Skoog, Physiol. Plant., 1962, 15, 473–497 CrossRef CAS.
- X. Li, A. Ahlman, H. Lindgren and L.-H. Zhu, Ind. Crops Prod., 2011, 33, 170–175, DOI:10.1016/j.indcrop.2010.10.004.
- A. Piovan, G. Cassina and R. Filippini, Biodiversity Conserv., 2011, 20, 359–371, DOI:10.1007/s10531-010-9949-z.
- B. G. Bowes, Protoplasma, 1976, 89, 185–188, DOI:10.1007/BF01279338.
- N. O. Pushkarova, PhD thesis, Institute of Cell Biology and Genetic Engineering, National Academy of Sciences of Ukraine, 2017.
- IUCN Red List of Threatened Species, http://www.iucnredlist.org, accessed December 2021 Search PubMed.
- M. Bilz, S. P. Kell, N. Maxted, and R. V. Lansdown European Red List of Vascular Plants, Publications Office of the European Union, Luxembourg, 2011, p. 142 Search PubMed.
- V. Radchuk, R. Radchuk, Ya. Pirko, R. Vankova, A. Gaudinova, V. Korkhovoy, A. Yemets, H. Weber, W. Weschke and Y. Blume, J. Exp. Bot., 2012, 63, 5497–5506, DOI:10.1093/jxb/ers200.
- H. M. Wang, T. Ketela, W. A. Keller, S. C. Gleddie and G. G. Brown, Plant Mol. Biol., 1995, 27, 801–807, DOI:10.1007/BF00020232.
- W. Youping and L. Peng, Euphytica, 1998, 101, 1–7, DOI:10.1023/A:1018384130269.
- E. Mietkiewska, J. M. Brost, E. M. Giblin, D. L. Barton and D. C. Taylor, Plant Biotechnol. J., 2007, 5, 636–645, DOI:10.1111/j.1467-7652.2007.00268.x.
- J. H. Mulder and H. D. Mastebroek, Euphytica, 1996, 89, 267–278, DOI:10.1007/BF00034615.
- R. J. Snowdon, Chromosome Res., 2007, 15, 85–95, DOI:10.1007/s10577-006-1105-y.
- A. Mohanty, B. Chrungu, N. Verma and K. R. Shivanna, Czech J. Genet. Plant Breed., 2009, 45, 117–122, DOI:10.17221/35/2009-CJGPB.
- S. I. Warwick and R. Gugel, Genet. Resour. Crop Evol., 2003, 50, 291–305, DOI:10.1007/s10722-004-2910-9.
- X. Z. Du, B. L. Huang, H. Guan, Z. Y. Li and B. Q. Huang, Genet. Mol. Res., 2014, 13, 6995–7005, DOI:10.4238/2014.March.26.2.
- J. Wen, J. X. Tu, Z. Y. Li, T. D. Fu, C. Z. Ma and J. X. Shen, Euphytica, 2008, 162, 81–89, DOI:10.1007/s10681-007-9566-4.
- S. Prakash, S. R. Bhat, C. F. Quiros, P. B. Kirti and V. L. Chopra, Plant Breed. Rev., 2009, 31, 21–187, DOI:10.1002/9780470593783.ch2.
- S. Hayta, M. A. Smedley, S. U. Demir, R. Blundell, A. Hinchliffe, N. Atkinson and W. A. Harwood, Plant Methods, 2019, 15, 121, DOI:10.1186/s13007-019-0503-z.
- X. Li, A. Ahlman, X. Yan, H. Lindgren and L. Zhu, Plant Cell, Tissue Organ Cult., 2010, 100, 149–156 CrossRef CAS.
- E. Mietkiewska, T. L. Hoffman, J. M. Brost, E. M. Giblin, D. L. Barton, T. Francis, Y. Zhang and D. C. Taylor, Mol. Breed., 2008, 22, 619627, DOI:10.1007/s11032-008-9204-4.
- S. Jeppson, K. Demski, A. S. Carlsson, L. H. Zhu, A. Banaś, S. Stymne and I. Lager, Front. Plant Sci., 2019, 10, 1442, DOI:10.3389/fpls.2019.01442.
- X. Qi, H. An, A. P. Ragsdale, T. E. Hall, R. N. Gutenkunst, J. Chris Pires and M. S. Barker, Mol. Ecol., 2017, 26, 3373–3388, DOI:10.1111/mec.14131.
- R. Guan, I. Lager, X. Li, S. Stymne and L. H. Zhu, Plant Biotechnol. J., 2014, 12, 193–203, DOI:10.1111/pbi.12128.
- J. Han, W. Lühs, K. Sonntag, U. Zähringer, D. S. Borchardt, F. P. Wolter, E. Heinz and M. Frentzen, Plant Mol. Biol., 2001, 46, 229–239, DOI:10.1023/A:1010665121980.
- X. Li, D. Mei, Q. Liu, J. Fan, S. Singh, A. Green, X. R. Zhou and L. H. Zhu, Plant Biotechnol. J., 2016, 14, 323–331, DOI:10.1111/pbi.12386.
- X. Li, R. Guan, J. Fan and L.-H. Zhu, Plant Cell Physiol., 2019, 60, 1274–1283, DOI:10.1093/pcp/pcz053.
- A. Giri and M. L. Narasu, Biotechnol. Adv., 2000, 18, 1–22, DOI:10.1016/s0734-9750(99)00016-6.
- R. K. Tiwari, M. Trivedi, Z. C. Guang, G. Q. Guo and G. C. Zheng, Biol. Plant., 2008, 52, 26–35, DOI:10.1007/s10535-008-0004-9.
- S. Ramachandra Rao and G. Ravishankar, Biotechnol. Adv., 2002, 20, 101–153, DOI:10.1016/s0734-9750(02)00007-1.
- E. Cequier-Sánchez, C. Rodríguez, R. Dorta-Guerra, A. G. Ravelo and R. Zarate, BMC Biotechnol., 2011, 11, 42, DOI:10.1186/1472-6750-11-42.
- E. Cequier-Sánchez, C. Rodríguez, A. G. Ravelo and R. Zárate, J. Agric. Food Chem., 2008, 56, 4297–4303, DOI:10.1021/jf073471e.
- B. Głab, T. Furmanek, M. Miklaszewska, A. Banas and A. Krolicka, Acta Physiol. Plant., 2013, 35, 2137–2145, DOI:10.1007/s11738-013-1250-9.
- M. Miklaszewska, A. Banaś and A. Królicka, Biotechnol. Bioeng., 2017, 114, 1275–1282, DOI:10.1002/bit.26234.
- T. K. Mohanta, T. Bashir, A. Hashem, E. F. Abh Allah and H. Bae, Genes, 2017, 8, 399, DOI:10.3390/genes8120399.
- A. Malzahn, L. Lowder and Y. Qi, Cell Biosci., 2017, 7, 21, DOI:10.1186/s13578-017-0148-4.
- U. Subedi, K. N. Jayawardhane, X. Pan, J. Ozga, G. Chen, N. A. Foroud and S. D. Singer, Lipids, 2020, 55, 495–512, DOI:10.1002/lipd.12249.
- J. J. Thelen and J. B. Ohlrogge, Metab. Eng., 2002, 4, 12–21, DOI:10.1006/mben.2001.0204.
- K. Zhang, L. Nie, Q. Cheng, Y. Yin, K. Chen, F. Qi and M. Li, Biotechnol. Biofuels, 2019, 12, 225, DOI:10.1186/s13068-019-1567-8.
- C. Morineau, Y. Bellec, F. Tellier, L. Gissot, Z. Kelemen, F. Nogué and J. D. Faure, Plant Biotechnol. J., 2017, 15, 729–739, DOI:10.1111/pbi.12671.
- W. Z. Jiang, I. M. Henry, P. G. Lynagh, L. Comai, E. B. Cahoon and D. P. Weeks, Plant Biotechnol. J., 2017, 15, 648–657, DOI:10.1111/pbi.12663.
- M. Yuan, J. Zhu, L. Gong, L. He, C. Lee, S. Han and G. He, BMC Biotechnol., 2019, 19, 24, DOI:10.1186/s12896-019-0516-8.
- M. McGinn, W. B. Phippen, R. Chopra, S. Bansal, B. A. Jarvis, M. E. Phippen and J. C. Sedbrook, Plant Biotechnol. J., 2019, 17, 776–788, DOI:10.1111/pbi.13014.
- J. A. Aznar-Moreno and T. P. Durrett, Plant Cell Physiol., 2017, 58, 1260–1267, DOI:10.1093/pcp/pcx058.
| This journal is © The Royal Society of Chemistry 2022 |