
Metabolite variations in the sera of HIV+ patients after an oral administration of effervescent glutamine and in comparison to non-HIV individuals by NMR†
Lucas Gelain
Martins
 *a,
Natália
Fregonesi
a,
Roberto Barbosa
Bazotte
b,
Jeane Eliete Laguila
Visentainer
c and
Ljubica
Tasic
*a
*a,
Natália
Fregonesi
a,
Roberto Barbosa
Bazotte
b,
Jeane Eliete Laguila
Visentainer
c and
Ljubica
Tasic
*a
aUNICAMP, Chemical Biology Laboratory, Department of Organic Chemistry, Institute of Chemistry, P. O. Box 6154, Campinas, São Paulo, Brazil. E-mail: lgmartins1984@gmail.com; ljubica@unicamp.br
bUEM, Department of Pharmacology and Therapeutics, Maringa, PR, Brazil
cUEM, Laboratory of Immunogenetics, Department of Basic Health Sciences, Maringa, PR, Brazil
First published on 10th September 2022
Abstract
It was demonstrated that effervescent glutamine supplementation in HIV+ individuals treated with antiretroviral therapy (ART) increased CD4+ T lymphocytes, decreased inflammation biomarkers, and brought health benefits. This pilot study aimed to explore serum metabolite variations in the HIV+ group under ART after 30 days of supplementation with glutamine, and in comparison to the matched HIV− group. The group of HIV+ showed lower levels of choline, creatine, pyruvate, glutamate, lysine, and tyrosine when compared to the HIV− group. Glucose, lipids, lactate, glutamine, phenylalanine, threonine, and phenylalanine/tyrosine were higher in HIV+ patients under long ART. Serum metabolome variations were shown to be consistent with the health improvements observed in the HIV+ group after effervescent glutamine supplementation, which might aid in ART in HIV+ individuals.
Introduction
HIV infection alters human cellular metabolism in many ways, favoring anabolic pathways, such as biosynthesis of nucleotides, phospholipids, cholesterol, amino acids, proteins, and glycoproteins, among others that are used for the assembly of newly synthesized viral particles.1–4 Therefore, lipid, protein, and carbohydrate metabolism are expected to change greatly.1 Additionally, antiretroviral therapy (ART) influences human metabolism by provoking side effects including metabolic syndrome which is characterized by increased abdominal fat, insulin resistance, type 2 diabetes, increased blood levels of free fatty acids, increased low-density lipoproteins (LDL), and very-low-density lipoproteins (VLDL), and, not rarely, premature aging.2,5,6Also, it has been demonstrated that the immune system and metabolism are intimately connected, and any alteration in one of them causes changes in the other.2,5 Infection with HIV causes depletion in CD4+ T lymphocytes, modifies nutrient demands, and disturbs the metabolic balance.4 It is also common to expect modified levels of glutamine, glutamate, and alanine–aspartate–glutamate in HIV+ individuals.7
Interesting metabolomic research on HIV/AIDS serum, and plasma, applying targeted or untargeted approaches3,7–11 had been published, with revealed changes in lipids, glucose, amino acids, choline, proteins, valine, and glutamine levels.12 Glutamine plays many important metabolic roles, as a precursor of glutathione, purines, and glutamate. As an indirect precursor of alpha-ketoglutarate,13 which is transferred to the mitochondria as the tricarboxylic acid cycle intermediate.
In our previous study,14 HIV+ individuals under long ART receiving glutamine supplementation for 30 days showed improvement suggestive of the benefits of this coadjuvant therapy. Surprisingly, no changes in the glutamine serum levels were observed, as opposed to the expectation, while improvement of CD4+ T lymphocytes and several other health parameters occurred. For this reason, by using the same cohort from our previous study, here we evaluated serum metabolome changes before and after glutamine supplementation, applying NMR-based metabolomics.
Results and discussion
HIV+ patients (P) versus healthy controls (C)
The impact of the viral infection on serum metabolites in HIV+ individuals under ART was investigated in comparison to the control group. 1H-NMR data were analyzed using supervised multivariate analyses. The best results were obtained in PLS-DA from T2-edited, as shown in Fig. 1 (accuracy: 0.980; R2: 0.999, and Q2: 0.699), and OPLS-DA, which is shown in Fig. 1C (permutation analysis R2Y: 0.998 for p < 0.05, and Q2: 0.712 for p < 0.05). There was one outlier (sample 16_2), which even when excluded did not influence the repeated PLS-DA model. Therefore, the model with this sample was analyzed further to avoid overfitting. The cluster analysis presented in the heatmap (Fig. 1D) also showed an excellent separation of groups, without any misclassification. The PLS-DA and OPLS-DA models for the WATERGATE and diffusion datasets are shown in the ESI† (Fig. S1).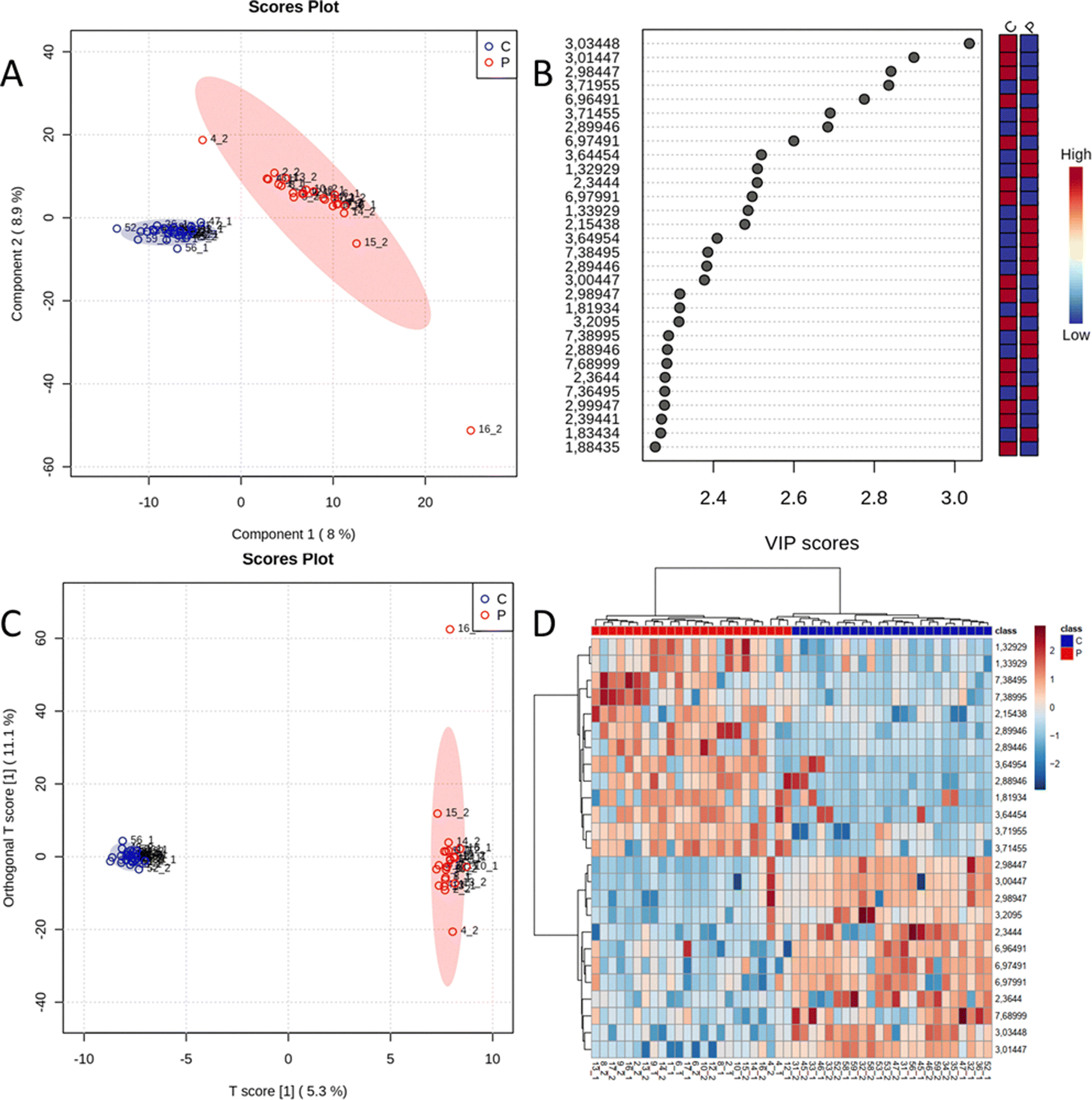 | ||
| Fig. 1 Multivariate analyses were performed on the serum 1H-NMR CPMG data of HIV+ patients treated with antiretroviral therapy (P) and HIV− individuals (C). (A) PLS-DA 2D scores (accuracy: 0.980, R2: 0.999, and Q2: 0.665). (B) PLS-DA VIP values. (C) OPLS-DA 2D scores. (D) Clustering results shown as a heatmap (distance measured using Euclidean and the clustering algorithm using Ward's method for 25 spectral bins/metabolites made by a t-test/ANOVA). | ||
The metabolites with VIP values higher than 2.2 (Fig. 1B) were analyzed with univariate analysis (t-test analysis, ESI,† Fig. S2, and Table S2), with boxplots illustated in Fig. 2. Details on the identification of metabolites are given in ESI† – Fig. S3 and S4, and Table S3. The ROC curve analysis (ESI,† Fig. S5) showed excellent AUC values (>0.888).
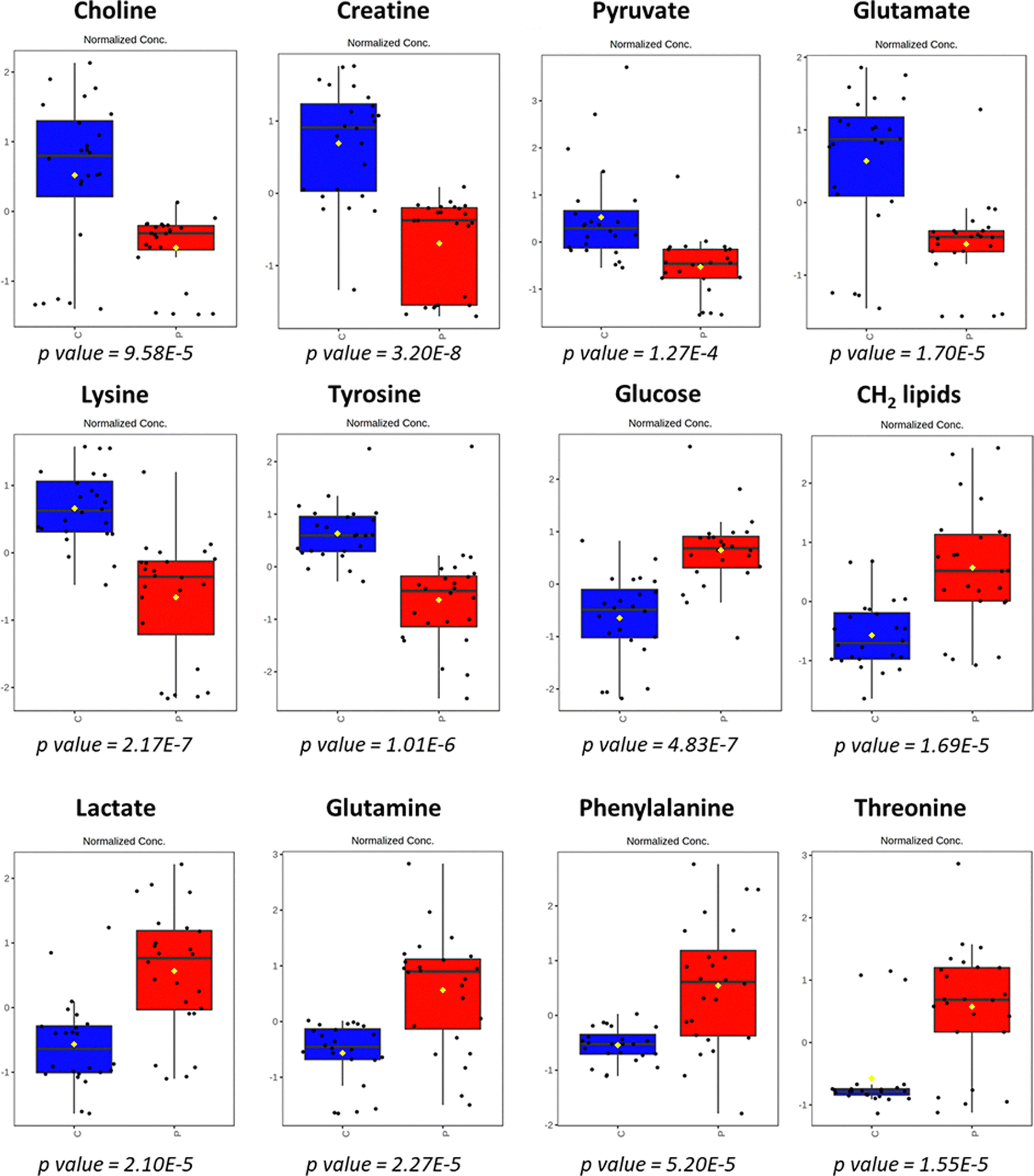 | ||
| Fig. 2 Boxplots of the main serum metabolites, which were found to be different between the HIV+ (P, red boxes) and HIV− individuals (C, blue boxes). Each box contains the metabolite variation according to a univariate t-test analysis. | ||
The results showed that HIV+ patients under ART presented lower levels of choline, creatine, pyruvate, glutamate, lysine, and tyrosine when compared to the control group. On the other hand, serum concentrations of glucose, lipids, lactate, glutamine, phenylalanine, and threonine were higher (p < 0.05) in HIV+ patients when compared with the control group. Once the metabolite variations due to the HIV infection were established, further analysis of metabolite variations within the same group of patients supplemented with effervescent glutamine was performed.
HIV before glutamine administration (BT) versus the same patients’ group after glutamine administration (AT)
Multivariate and univariate analyses were applied to the same group of patients before (BT) and after treatment (AT) with effervescent glutamine. The PLS-DA and OPLS-DA (Fig. 3) distinguished the groups with good accuracy and linear regression fitting (R2). However, the model was not predictive (low Q2), and thus the VIP scores were not used for biomarker analysis. Metabolite changes were evaluated using univariate t-test analysis (ESI,† Fig. S6 and Table S4), and the biomarkers that showed statistically significant variations (p-value < 0.05) are shown in Fig. 4. Finally, the univariate analysis of the ROC curve was performed (ESI,† Fig. S7) for each of the metabolites. | ||
| Fig. 3 Multivariate analyses were performed on the serum 1H-NMR CPMG data of HIV+ patients treated with ART before (BT) and after treatment (AT) with effervescent glutamine (EG). (A) PLS-DA 2D scores (accuracy: 0.564, R2: 0.964, and Q2: −0.085). (B) OPLS-DA 2D scores. | ||
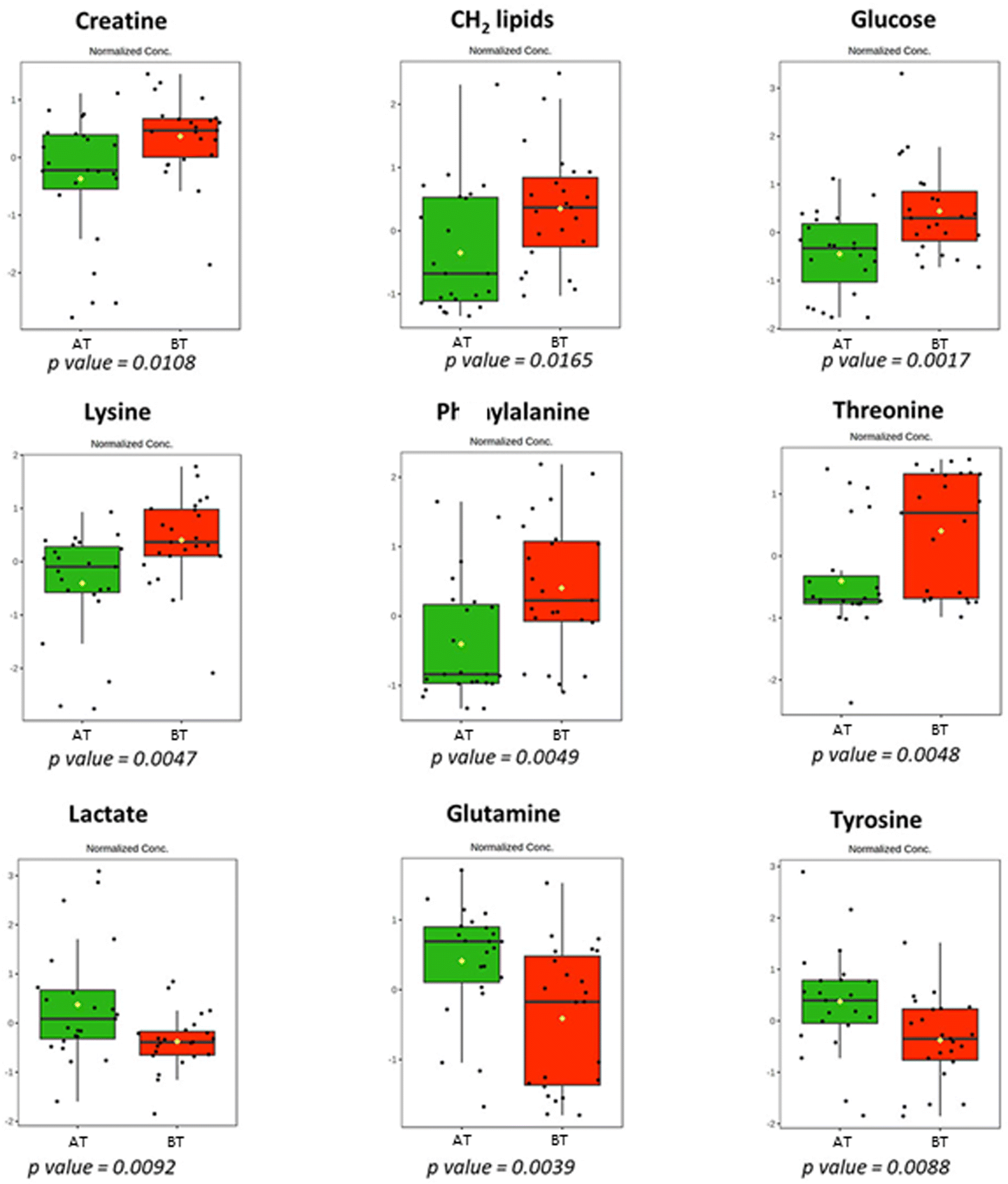 | ||
| Fig. 4 Boxplots showing discriminatory metabolites in HIV+ patients before (BT, red box) and after (AT, green box) treatment with EG. Each box contains the metabolite variation according to a univariate t-test analysis. | ||
The univariate t-test and ROC results enabled us to select nine biomarkers that differ between AT and BT groups. The EG administration reduced the serum concentrations of creatine, lipids, glucose, lysine, phenylalanine, and threonine. The AT group showed increased serum concentrations of lactate, glutamine, and tyrosine.
As already published, the AT group showed an increase of blood CD4+ T cells,14 but the values of CD8+ T cells and CD48+ T cells remained unchanged. T cells are very important for the immune response during infection by the specific T-cell receptors that mediate antigen signaling, and lead to increased T cell uptake of phenylalanine, leucine, and glutamine,17 which is consistent with the decreased phenylalanine concentration in serum (as can be seen from Fig. 4).18 It was reported that HIV+ patients have a higher ratio of phenylalanine/tyrosine than healthy individuals,19 and our results are in agreement which can be verified from the metabolite box plot in Fig. 2. As phenylalanine is a tyrosine precursor, which in turn is a precursor of neurotransmitters such as dopamine, norepinephrine, and epinephrine, a high phenylalanine/tyrosine ratio means low neurotransmitter synthesis, a phenomenon related to disease-associated neuropsychiatric symptoms, which are common in HIV+ patients.20 Interestingly, dietary supplementation with EG caused a decrease in phenylalanine and an increase in the serum tyrosine of the HIV+ group, as shown in Fig. 4, which results in a decrease in phenylalanine/tyrosine.
Although a slight increase in serum glutamine was observed after treatment (Fig. 4), it is evident that a large part of the administered glutamine was consumed by CD4+ T lymphocytes. The ASCT2 amino acid transport mediates the entry of glutamine into the cell resulting in immunostimulant activity.21
Glutamine also presents interactions on the mammalian target of rapamycin complex 1 (mTORC1), activating, through indirect mechanisms, cell metabolism and differentiation. The mechanism by which glutamine shows beneficial effects on the immune system might involve its role as the primary fuel source for immune cells. In addition, glutamine was well tolerated as no changes were observed in the serum parameters of nephrotoxicity (creatinine), hepatotoxicity (alanine aminotransferase and aspartate aminotransferase), and amino acid catabolism (urea).14
In addition to the increased CD4+ T cells, which are important for improving the immune response in HIV+ patients, other health improvements were observed. There was a marked decrease (P < 0.01) in the serum levels of several parameters of inflammation (C-reactive protein, interferon γ inducible protein, macrophage inflammatory protein-1β, and RANTES) after effervescent glutamine supplementation.14 In agreement with these results, glutamine acted not only as a metabolic fuel, but also as a precursor of glutathione and regulated the expression of genes that encode inflammatory cytokines. For example, the immunomodulatory effects of glutamine are mediated by the expression of FOXP3, a transcription factor that also modulates CD4+ T cell proliferation and inflammatory response.
The high energy demand of T cells justifies the consumption of glucose through aerobic glycolysis, which culminates in lactate production. Decreased glucose and increased lactate were observed after glutamine administration in the HIV+ group (Fig. 4). When CD4+ T lymphocytes are under aerobic glycolysis conditions, the tricarboxylic acid cycle needs a replacement, which comes from glutaminolysis.17,22,23 The higher serum glutamine levels and lower serum glutamate levels suggest a decreased metabolism of glutamine in the HIV+ group.
Although creatine and threonine metabolism pathways are altered in Jurkat T-cells grown in the presence of endotoxin-free recombinant HIV−1 clade B Tat protein, which is released by infected cells, these amino acids are not recognized as important biomarkers of HIV infection.24
Experimental section
Cohort selection and patient treatment
The clinical procedures were conducted according to the ethical standards established in the Declaration of Helsinki. The Maringá State University Ethics Committee (COPEP 1.166.674) and the Brazilian Clinical Trials Register (RBR843tnq) approved the protocol of the study.All patients signed the Informed Consent Form that includes the following statement: “The collected blood will be stored in the form of serum or plasma. If there is a need for additional dosages in the serum or plasma, these will be performed as long as they do not interfere with your privacy. We also remember that the information obtained will be used only for research purposes and will be treated with confidentiality to preserve your privacy”. Therefore, sera from all HIV+ patients under ART14,15 were used. Data concerning inclusion and exclusion criteria, age, gender, medical history, educational level, time with AIDS, therapeutic approach for ART, lifestyle, biochemical, toxicological, hematological, and inflammatory parameters can be found in our previous studies.14,15
The patients enrolled in this study were from the Public Service Health Care for Sexually Transmitted Diseases/AIDS Center of Maringá City, Paraná, Brazil, in the period of 2012–2015. Our initial cohort included 2534 HIV+ patients treated with ART.
Patients who were transferred, who abandoned treatment, and who had not been evaluated for viral load and CD4+ T lymphocyte counts were excluded. To differentiate the improvement caused by the administration of glutamine and not due to ART, we selected patients who had stable CD4+ T cell counts (<500 cells mm−3), who were currently were receiving an unchanged ART for at least one year before starting the clinical investigation, and who took the supplementation on a regular basis for 30 days. For these reasons, the number of individuals studied dropped from 2534 to just 12. The HIV− individuals were carefully selected and matched by age and gender with the HIV+ patients. Therefore, the cohort selected for this study comprises 12 patients living with HIV treated with ART (in duplicate), the same 12 patients after 30 days of ART plus effervescent glutamine treatment (also in duplicate), and a group for comparison studies, named healthy donors (12 individuals), who were also analyzed in duplicate.
Because glutamine (Ajinomoto North America, NC, USA, lot 140165K03B) is only slightly soluble in water, a palatable effervescent glutamine formulation prepared in a microparticulate system that increased its solubility in water was prepared. This novel glutamine formulation developed by our research group16 was orally administered to the 12 HIV/AIDS individuals included in this study. The patients were required to dissolve the effervescent glutamine formulation (supplied in sachets) in a glass of water (240 mL) immediately before oral ingestion (12.4 g), once a day, after lunch, or after dinner for 30 days.
Serum samples and 1H-NMR analyses
Serum samples were obtained from the 12 HIV+ patients before (day 0) and after glutamine (day 31) administration, and from 12 healthy donors. The aliquots (250 μL) were kept in cryovials at −80 °C. Prior to analysis, the samples were thawed on ice, mixed with 250 μL of deuterium oxide (D2O 99.9%, Cambridge Isotope Laboratories, Inc., Andover, USA) at room temperature, centrifuged at 12![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 × g for 2min at 4 °C, and transferred into 5 mm NMR tubes. High-resolution 1H-NMR spectra with water signal elimination (WATERGATE), 1H-NMR spectra with T2 filter (cpmgpr1d), and 1H-NMR spectra edited by diffusion (stebpgp1s191d) were acquired on a Bruker AVANCE III 600 MHz spectrometer using an inverse triple-core probe (TBI) at 25 °C. Total correlation spectroscopy (TOCSY) spectral data were recorded using the mlevphpr pulse sequence. 1D and 2D NMR data, such as chemical shifts, coupling constants, the multiplicity of peaks, and the human metabolomic database (HMDB), were used for metabolite assignments.
000 × g for 2min at 4 °C, and transferred into 5 mm NMR tubes. High-resolution 1H-NMR spectra with water signal elimination (WATERGATE), 1H-NMR spectra with T2 filter (cpmgpr1d), and 1H-NMR spectra edited by diffusion (stebpgp1s191d) were acquired on a Bruker AVANCE III 600 MHz spectrometer using an inverse triple-core probe (TBI) at 25 °C. Total correlation spectroscopy (TOCSY) spectral data were recorded using the mlevphpr pulse sequence. 1D and 2D NMR data, such as chemical shifts, coupling constants, the multiplicity of peaks, and the human metabolomic database (HMDB), were used for metabolite assignments.
Statistical analysis
The serum 1H-NMR spectra (WATERGATE, CPMG, or edited by diffusion) were baseline- and phase-corrected, aligned, and referenced (lactate signal, 3H, δ 1.33, d, J = 7.0 Hz) using MestreNova Inc. Software. The HIV+ individuals before glutamine administration (12 patients in duplicate, n = 24, named “P”) were compared with the healthy control donors (12 individuals in duplicate, n = 24, named “C”). Then, spectra from the HIV+ group before glutamine administration (n = 24, now named “BT” from before the treatment) were compared to ones recorded after 30 days of glutamine administration (n = 24, now named “AT” from after the treatment). The spectra were normalized by mean centering, the water peak was excluded, binned (0.005 ppm), and exported as data in the .csv format for the chemometrics analysis. Chemometrics were done using the MetaboAnalyst 5.0 software platform (https://www.metaboanalyst.ca/). The following analyses were performed: partial least squares discriminant analysis (PLS-DA) with leave-one-out cross-validation, orthogonal projections to latent structures discriminant analysis (oPLS-DA), heatmap clustering analysis, univariate t-test analysis, and the receiver operating characteristic (ROC) curve.Conclusions
The main limitation of this study is the small sample size because of difficulties in the follow-up and treatment results of the HIV+ patients. Nevertheless, the carefully selected group underwent the same ART and effervescent glutamine treatments, which validates the observed variations in metabolites. Metabonomics by 1H-NMR was applied successfully to the follow-up of HIV+ patients.Data availability
This data is available at the NIH Common Fund's National Metabolomics Data Repository (NMDR) website, the Metabolomics workbench, https://www.metabolomicsworkbench.org where it has been assigned Project ID PR001351. The data can be accessed directly via it's Project https://doi.org/10.21228/M83699.Author contributions
R. B. B., L. T. and L. G. M. conceived and designed the experiments. R. B. B. and J. E. L. V. selected cases and controls and took part in the clinical evaluation of patients. L. G. M. and N. F. carried out the experiments and analyzed and interpreted the data. L. G. M. and L. T. wrote the paper. All authors reviewed and approved the manuscript.Conflicts of interest
There are no conflicts to declare.Acknowledgements
The authors would like to acknowledge the funding from the Sao Paulo Research Foundation, FAPESP grants #2018/24069-3 and #2018/06510-4, and INCTBio. PPSUS, Brazil, Protocol 41645.433.30361.12092013, is also kindly acknowledged.Notes and references
- E. L. Sanchez and M. Lagunoff, Viral activation of cellular metabolism, Virology, 2015, 479–480, 609–618, DOI:10.1016/j.virol.2015.02.038.
- S. U. Munshi, B. B. Rewari, N. S. Bhavesh and S. Jameel, Nuclear magnetic resonance-based profiling of biofluids reveals metabolic dysregulation in HIV-infected persons and those on antiretroviral therapy, PLoS One, 2013, 8, e64298, DOI:10.1371/journal.pone.0064298.
- Y. Ding, H. Lin, X. Chen, B. Zhu, X. Xu, X. Xu, W. Shen, M. Gao and N. He, Comprehensive metabolomics profiling reveals common metabolic alterations underlying the four major non-communicable diseases in treated HIV infection, EBioMedicine, 2021, 71, 103548, DOI:10.1016/j.ebiom.2021.103548.
- R. D. Michalek, V. A. Gerriets, S. R. Jacobs, A. N. Macintyre, N. J. Maclver, E. F. Mason, S. A. MasonSullivan, A. G. Nichols and J. C. Rathmell, Cutting edge: Distinct glycolytic and lipid oxidative metabolic programs are essential for effector and regulatory CD4+ T cell subsets, J. Immunol., 2011, 186, 3299–3303, DOI:10.4049/jimmunol.1003613.
- H. L. Yeoh, A. C. Cheng, A. C. Cherry, J. M. Weir, P. J. Meikle, J. F. Hoy, S. M. Crowe and C. S. Palmer, Immunometabolic and lipidomic markers associated with the frailty index and quality of life in aging HIV+ men on antiretroviral therapy, EBioMedicine, 2017, 22, 112–121, DOI:10.1016/j.ebiom.2017.07.015.
- S. U. Kaur, B. F. Oyeyemi, A. Shet, B. P. Gopalan, D. Himanshu, N. S. Bhavesh and R. Tandom, Plasma metabolomic study in perinatally HIV-infected children using 1H NMR spectroscopy reveals perturbed metabolites that sustain during therapy, PLoS One, 2020, 15, e0238316, DOI:10.1371/journal.pone.0238316.
- L. J. Sitole, F. Tugizimana and D. Meyer, Multi-platform metabonomics unravel amino acids as markers of HIV/combination antiretroviral therapy-induced oxidative stress, J. Pharm. Biomed. Anal., 2019, 176, 112796, DOI:10.1016/j.jpba.2019.112796.
- R. Hewer, J. Vorster, F. E. Steffens and D. Meyer, Applying biofluid 1H NMR-based metabolomic techniques to distinguish between HIV-1 positive/AIDS patients on antiretroviral treatment and HIV-1 negative individuals, J. Pharm. Biomed. Anal., 2006, 41, 1442–1446, DOI:10.1016/j.jpba.2006.03.006.
- A. C. Justice, K. M. Erlandson, P. W. Hunt, A. Landay, P. Miotti and R. P. Tracy, Can biomarkers advance HIV research and care in the antiretroviral therapy era?, J. Infect. Dis., 2018, 217, 521–528, DOI:10.1093/infdis/jix586.
- A. Williams, K. Kgoadi, F. Steffens, P. Steenkamp and D. Meyer, UPLC-MS Metabonomics reveals perturbed metabolites in HIV-infected sera, Curr. Metabolomics, 2014, 2, 37–52, DOI:10.2174/2213235x02666140214201251.
- C. Philippeos, F. E. Steffens and D. Meyer, Comparative 1H NMR-based metabonomic analysis of HIV-1 sera, J. Biomol. NMR, 2009, 44, 127–137, DOI:10.1007/s10858-009-9329-8.
- I. Clerc, D. A. Moussa, Z. Vahlas, S. Tardito, L. Oburoglu, T. J. Hope, M. Sitbon, V. Dardalhon, C. Mongellaz and N. Taylor, Entry of glucose- and glutamine-derived carbons into the citric acid cycle supports early steps of HIV-1 infection in CD4 T cells, Nat. Metab., 2019, 1, 717–730, DOI:10.1038/s42255-019-0084-1.
- F. J. Vázquez-Santiago, R. J. Noel, J. T. Porter and V. Rivera-Amill, Glutamate metabolism and HIV-associated neurocognitive disorders, J. Neurovirol., 2014, 20, 315–331, DOI:10.1007/s13365-014-0258-2.
- T. Krupek, B. J. W. Ferrari, M. A. R. C. P. Silva, C. R. Schamber, D. A. Bertolini, M. L. Bruschi, I. T. S. Previdelli, O. C. N. Pereira, J. R. C. Dias, R. Curi and R. B. Bazotte, Effervescent glutamine formulation improves the beneficial effects of antiretroviral therapies on immune function in HIV/AIDS carrier patients, J. Med. Food, 2020, 23, 5, DOI:10.1089/jmf.2019.0129.
- G. Godoy, M. M. Antunes, M. A. R. C. P. da Silva, I. L. Fernandes, J. V. Visentainer, J. L. Visentainer, R. Curi and R. B. Bazotte, Decreased docosahexaenoic acid levels in serum of HIV carrier patients, J. Med. Food, 2021, 24(6), 670–673, DOI:10.1089/jmf.2020.0119.
- B. J. Ferrari, B. L. Pelegrini, J. B. Da Silva, O. C. N. Pereira, M. M. S. Lima, M. L. Bruschi and R. B. Bazotte, Formulation and in vivo study of the solid effervescent system as a new strategy for oral glutamine delivery, J. Drug Delivery Sci. Technol., 2021, 63, 102516, DOI:10.1016/j.jddst.2021.102516.
- J. Wei, J. Raynor, T. L. M. Nguyen and H. Chi, Nutrient and metabolic sensing in T cell responses, Front. Immunol., 2017, 8, 247, DOI:10.3389/fimmu.2017.00247.
- F. Castellano and V. Molinier-Frenkel, Control of T-Cell Activation and Signaling by Amino-Acid Catabolizing Enzymes, Front. Cell Dev. Biol., 2020, 8, 613416, DOI:10.3389/fcell.2020.613416.
- G. P. Innocenti, L. Santinelli, L. Laghi, C. Borrazzo, C. Pinacchio, M. Fratino, L. Celani, E. N. Cavallari, C. Scagnolari, F. Frasca, G. Antonelli, C. M. Mastroianni, G. d’Ettorre and G. Ceccarelli, Modulation of phenylalanine and tyrosine metabolism in HIV-1 infected patients with neurocognitive impairment: Results from a clinical trial, Metabolites, 2020, 10, 274, DOI:10.3390/metabo10070274.
- J. M. Gostner, K. Becker, K. Kurz and D. Fuchs, Disturbed amino acid metabolism in HIV: Association with neuropsychiatric symptoms, Front. Psychiatry, 2015, 6, 97, DOI:10.3389/fpsyt.2015.00097.
- M. Villa Nova, B. A. Ratti, L. S. Herculano, P. R. S. Bittencourt, C. R. Novello, R. B. Bazotte, S. O. S. Lautenschlager and M. L. Bruschi, Design of composite microparticle systems based on pectin and waste material of propolis for modified l-alanyl-l-glutamine release and with immunostimulant activity, Pharm. Dev. Technol., 2019, 24(1), 12–23, DOI:10.1080/10837450.2017.
- M. C. Poffenberger and R. G. Jones, Amino acids fuel T cell-mediated inflammation, Immunity, 2014, 40, 635–637, DOI:10.1016/j.immuni.2014.04.015.
- C. S. Palmer, M. Ostrowski, B. Balderson, N. Christian and S. M. Crowe, Glucose metabolism regulates T cell activation, differentiation, and functions, Front. Immunol., 2015, 6, 1, DOI:10.3389/fimmu.2015.00001.
- W. Liao, G. Tan, Z. Zhu, Q. Chen, Z. Lou, X. Dong, W. Zhang, W. Pan and Y. Chai, Combined metabonomic and quantitative real-time PCR analyses reveal systems metabolic changes in Jurkat T-cells treated with HIV-1 Tat protein, J. Proteome Res., 2012, 11, 5109–5123, DOI:10.1021/pr300173c.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d2mo00097k |
| This journal is © The Royal Society of Chemistry 2023 |