
The plasma phospholipidome of the bottlenose dolphin (Tursiops truncatus) is modulated by both sex and developmental stage†
João P.
Monteiro‡
 *ab,
Helena B.
Ferreira
ab,
Tânia
Melo
ab,
Carla
Flanagan
c,
Nuno
Urbani
c,
João
Neves
c,
Pedro
Domingues
ab and
M. Rosário
Domingues
ab
*ab,
Helena B.
Ferreira
ab,
Tânia
Melo
ab,
Carla
Flanagan
c,
Nuno
Urbani
c,
João
Neves
c,
Pedro
Domingues
ab and
M. Rosário
Domingues
ab
aMass Spectrometry Centre, LAQV-REQUIMTE, Department of Chemistry, University of Aveiro, Aveiro, Portugal. E-mail: jpspmonteiro@yahoo.com; Tel: +351 963261356
bCESAM-Centre for Environmental and Marine Studies, Department of Chemistry, University of Aveiro, Aveiro, Portugal
cZoomarine, Guia, Albufeira, Portugal
First published on 24th October 2022
Abstract
Lipidomics represent a valid complementary tool to the biochemical analysis of plasma in humans. However, in cetaceans, these tools have been unexplored. Here, we evaluated how the plasma lipid composition of Tursiops truncatus is modulated by developmental stage and sex, aiming at a potential use of lipidomics in integrated strategies to monitor cetacean health. We characterized the fatty acid profile and detected a total of 26 fatty acids in T. truncatus plasma. The most abundant fatty acids were palmitic acid (C16:0), stearic acid (C18:0) and oleic acid (C18:1n-9). Interestingly, there are consistent differences between the fatty acid profile of mature female and mature male specimens. Phospholipidome analysis identified 320 different lipid species belonging to phosphatidylcholine (PC, 105 lipid species), lysophosphatidylcholine (42), phosphatidylethanolamine (PE, 67), lysophosphatidylethanolamine (18), phosphatidylglycerol (14), lysophosphatidylglycerol (8), phosphatidylinositol (14), lysophosphatidylinositol (2), phosphatidylserine (3), sphingomyelin (45) and ceramides (2) classes. The statistical analysis of the phospholipidome showed that its composition allows discriminating mature animals between sexes and mature males from immature males. Notably, discrimination between sexes is mainly determined by the contents of PE plasmalogens and lysophospholipids (LPC and LPE), while the differences between mature and immature male animals were mainly determined by the levels of PC lipids. This is the first time that a correlation between developmental stage and sex and the lipid composition of the plasma has been established in cetaceans. Being able to discern between age and sex-related changes is an encouraging step towards using these tools to also detect differences related to disease/dysfunction processes.
Introduction
The bottlenose dolphin (Tursiops truncatus), is undoubtedly the best characterized and studied cetacean. They are the most common cetacean species maintained by humans around the world.1 Bottlenose dolphins adapt to a wide variety of marine and estuarine habitats, roaming most warm temperate and tropical seas, with brief incursions into rivers in some places.2 Their diet is mainly based on fish and/or squid, with a preference for sciaenids, scombrids, and mugilids, depending on the specific region/habitat they inhabit.3As adults, bottlenose dolphins exhibit a light grey to black colour dorsally and laterally and a lighter belly, measuring 2.5–3.8 meters in length.4 Females can live up to 67 years while males can live up to 52 years.2 Female bottlenose dolphins generally reach their sexual and physical maturity before males, between 5–13 years old, while males should reach this stage of development between 9–14 years old.2 For this work, we used the upper values of these intervals to set the age limit between mature and immature individuals for females and males.
As with humans, biochemical analyses of blood or plasma are also the most commonly used means of monitoring the health of managed cetaceans.5–7 Most of the available knowledge on blood analytes and plasma parameters/characteristics in cetaceans has been obtained from animals under human care. In fact, after diligent conditioning procedures including positive reinforcement, blood collection from these animals is as comfortable as possible. The ability to draw blood from managed animals has secured the resources to perform many studies, promoting significant advances in the elucidation of the idiosyncratic metabolism and physiology of these animals, also providing the baselines for the detection of dysfunction/disease. Circulating lipids in plasma/serum are, in fact, normally analysed for medical evaluation, trying to discern physiological and metabolic variations that are characteristic of pathological contexts,8,9 as in the case of cholesterol and triglyceride, normally linked to cardiovascular risk. With the emergence of new technical solutions and the development of lipidomics tools, numerous studies have demonstrated correlations between disease and alterations in plasma lipids, proposing specific lipid biomarkers of health and disease conditions.9–17 It is therefore foreseeable that corresponding tools can also be used for the benefit of cetacean species. Previously available studies have already set some intervals between which certain parameters (transferrin saturation) and analyte/metabolites (creatinine, glucose, insulin, triglycerides, cholesterol, free fatty acids, ceramides, adiponectin) take place in the plasma of T. truncatus in particular.7,18–20 Some of these parameters are currently being used to monitor dolphin health conditions. For instance, chronic elevations in triglycerides and LDL cholesterol levels have been associated with disease conditions in these animals.21 The study of lipid content and understanding of lipid metabolism is also of particular importance because, as most marine mammals, they feed on preys rich in lipids and proteins and almost devoid of carbohydrates.22 Furthermore, marine mammals were proposed to present a unique lipid metabolism,22–26 with a particular triacylglycerol metabolism24 and a peculiar glucose metabolism and fat-depot signalling functions.27 Interestingly, managed cetaceans were proposed as viable, interesting potential models able to mimic aspects of human pathophysiology,28 without some of the limitations imposed by the use short-lived animal models,29 which may not represent adequate models to fully appraise diseases or comorbidities normally associated with aging or for which age is a risk factor. Moreover, cetaceans, and dolphins in particular, do present manifestations of chronic diseases and comorbidities with a frame comparable to human conditions, such as chronic inflammation,29 cardiovascular disease-associated manifestations,30,31 non-alcoholic fatty liver disease,32,33 diabetes,34–36 and even histological lesions mimicking Alzheimer's disease,37,38 which do not naturally occur in the case short-lived mammal models.
Several studies have reported metabolic differences related to sex and age in in T. Truncatus. Both energy intake and total energy expenditure were shown to decrease with age, while fat mass was shown to increase.39,40 Important hormonal changes were also described, with thyroid hormones decreasing with age and greatly varying with sex, which may anticipate an influence in important physiological regulatory processes.41 Also, adrenal hormones were reported to vary with sex, age and other factors in T. Truncatus.42–44 Moreover, several other studies reported the impact of age and sex in the haematological parameters.45–48 Therefore, we anticipated that the unique lipid metabolism of these cetaceans and plasma lipid composition, could also be modulated by both sex and developmental stage. In a previous work, we characterized the lipid content in the plasma of mature male specimens of T. truncatus using a lipidomics approach.49 Another previous study had already probed the feasibility of using lipidomics plasma analyses to monitor the health of managed cetaceans but in this case beluga whales (Delphinapterus leucas),50 while some other work used lipidomics tools to characterize dolphin (T. truncatus)51 or whale (Orcinus orca and Megaptera novaeangliae)52 blubber.
Both studies highlighted how informative plasma lipid profiling can be, while also unveiling the potential of this approach to be part of diagnostic integrative efforts, along with routine plasma biochemical analyses.
Therefore, in this work, we investigated how the plasma phospholipidome of T. truncatus can be modulated by factors such as sex and developmental stage using state-of-the-art GC-MS (gas chromatography – mass spectrometry) and LC-MS (liquid chromatography – mass spectrometry) lipidomic approaches. On one hand, we believe that this work represents a privileged inside look into the factors that may regulate lipid metabolism in this species. On the other hand, this study also provides baselines for lipid occurrence in plasma, from where variations elicited by disease/dysfunction may be appraised. Therefore, this work represents an important step towards the further implementation of lipidomics tools and lipid biomarkers to monitor the health status of T. truncatus. This integration of lipidomics tools into cetacean health monitoring should result in more robust and thorough procedures, and the low technical detection limits should ensure preventive and timely detection advantages.
Experimental
Experimental ethics
This study was conducted in accordance with the guidelines for animal experiments.53 As part of a preventive medicine program implemented by the Zoomarine Veterinary Department, monitoring the health of dolphins includes routine monthly analysis of a blood sample. At Zoomarine Portugal, this preventive medicine program is possible thanks to the many husbandry behaviours carried out voluntarily, allowing the monitoring of the dolphin′s health to be routine and stress-free. The dolphins were trained using operant conditioning with positive reinforcement, for blood collection from the ventral side of their tail fluke. Prior to the beginning of the study, the project was evaluated by the Zoomarine's Science Committee for its feasibility and possible welfare implications, having been accepted with no restrictions (although no written recommendation was issued as this is an informal advisory body). Since the collection of samples was part of the on-going preventive medicine plan, no new samples were needed. As so, it had tacit approval by the competent national institutions (ICNF and DGAV).Samples
Plasma samples were obtained as previously described [22]. Briefly, the 23 dolphins included in this study (14 males; 9 females, two lactating: Dolphins 6 and 8; Table 1) were housed in an outside lagoon complex of the Zoomarine facilities with 9195 m3 of natural seawater. The dolphins’ diet consisted of thawed frozen fish of the following species in varying proportions: mackerel (Scomber japonicus), herring (Clupea harengus), capelin (Mallotus villosus), blue whiting (Micromesistius poutassou), horse mackerel (Trachurus trachurus), sprat (Sprattus sprattus), smelt (Atherina boyeri), sardine (Sardina pilchardus), pout (Trisopterus esmarkii) e sandeel (Ammodytes tobianus) and squid (Loligo vulgaris). In general, the same fish offer is available to all individuals regardless age or sex. Blood samples were collected in SARSTEDT S-Monovette 2,7 mL K3E tubes with SARSTEDT Safety-Multifly®21G tube 200 m butterfly needle (Nümbrecht, Germany). The samples were immediately centrifuged at 5000 rpm for 5 min (Pro-vet PCVS, Centurion Scientific, West Sussex, United Kingdom) and the plasma was transferred to 1.5 mL micro-tubes and refrigerated. The samples were stored at −80 °C until the analyses were carried out.| Sex | Age | Length (cm) | Experimental group | |
|---|---|---|---|---|
| FI: female immature (N = 3); FM: female Ma 1FI: female immature (N = 3); FM: female mature (N = 6); MI: male immature (N = 6); MM: male mature (N = 8). | ||||
| Dolphin 1 | Female | 3 | 229 | FI |
| Dolphin 2 | Female | 5 | 240 | FI |
| Dolphin 3 | Female | 9 | 223 | FI |
| Dolphin 4 | Female | 13 | 253 | FM |
| Dolphin 5 | Female | 14 | 243 | FM |
| Dolphin 6 | Female | 21 | 250 | FM |
| Dolphin 7 | Female | 22 | 252 | FM |
| Dolphin 8 | Female | 24 | 242 | FM |
| Dolphin 9 | Female | 42 | 257 | FM |
| Dolphin 10 | Male | 3 | 230 | MI |
| Dolphin 11 | Male | 6 | 226 | MI |
| Dolphin 12 | Male | 6 | 235 | MI |
| Dolphin 13 | Male | 8 | 238 | MI |
| Dolphin 14 | Male | 10 | 238 | MI |
| Dolphin 15 | Male | 11 | 250 | MI |
| Dolphin 16 | Male | 15 | 259 | MM |
| Dolphin 17 | Male | 16 | 258 | MM |
| Dolphin 18 | Male | 17 | 246 | MM |
| Dolphin 19 | Male | 20 | 282 | MM |
| Dolphin 20 | Male | 24 | 245 | MM |
| Dolphin 21 | Male | 24 | 249 | MM |
| Dolphin 22 | Male | 25 | 272 | MM |
| Dolphin 23 | Male | 27 | 268 | MM |
Chemicals
All solvents used were of HPLC-MS (high performance liquid chromatography – mass spectrometry) grade. Chemicals were purchased from Sigma-Aldrich (Steinheim, Germany) and had a purity higher than 95%. Milli-Q water (Synergy®, Millipore Corporation, Billerica, MA, USA) was used. Phospholipid internal standards were obtained from Avanti Polar Lipids (Alabaster, USA). In GC-MS analysis, the 37 Component FAME Mix from Supelco (Sigma Aldrich, St. Louis, MO, USA) was used as methyl ester of fatty acids (FAME) standard mix, while internal standard methyl heptadecanoate (≥99% purity) was purchased from Sigma (St. Louis, MO, USA).Lipid extraction
The polar lipid fractions from plasma samples were obtained by solid-phase extraction (SPE). The procedure required three different eluents used consecutively. The first eluent consisted of 1% of formic acid in acetonitrile, the second was (pure) acetonitrile, and the third and final eluent consisted of 5% of ammonium hydroxide in acetonitrile. First, 100 μL of plasma were mixed with 900 μL of the first eluent, vortexed (Vortex Labinco L46) for 30 seconds and centrifuged (Mixtasel, Model 540 P Selecta) at 626 × g for 5 min. The resulting supernatant was transferred to a Hybrid SPE-PL column (SUPELCO, lot 4497303) already placed in a vacuum system Visiprep SPE Vacuum Manifold (Supelco, Bellefonte, PA). After elution of almost all of the supernatant, the columns were washed with 1 mL of the second eluent, followed by 1 mL of the first eluent. At this point, the collection tubes were replaced with new ones and the polar lipid fraction retained on the Hybrid SPE-PL columns was eluted with two consecutive 1 mL aliquots of the third eluent. The flow-through was collected, dried under a stream of nitrogen and stored at −80 °C.Phospholipid quantification
Phospholipids in plasma polar lipid extracts were quantified by an adaptation of the Bartlett and Lewis method.54,55 Samples were dissolved in 200 mL dichloromethane and 10 μL aliquots (in duplicates) were transferred to acid-washed glass tubes and then dried under a nitrogen stream. A volume of 125 μL of perchloric acid (70%, m V−1) was added to each dried extract-containing tube. Tubes were incubated for 60 min at 180 °C in a steel heating block and after hydrolysis, 825 μL of water, 125 μL of ammonium molybdate (2.5%, m V−1) and 125 μL of ascorbic acid (10%, m V−1) were added to each tube, which were then maintained at 100 °C for 10 min in a water bath. A calibration curve was produced using phosphorous standards with concentrations ranging from 0.1 to 2.0 μg of (standard solution of NaH2PO4. 2H2O). Absorbance was measured in a microplate UV-Vis spectrophotometer Thermo Scientific Multiskan Go at 797 nm.Gas chromatography-mass spectrometry (GC-MS)
Esterified fatty acids in plasma lipid extracts were analyzed after transmethylation.56 Aliquots containing 10 μg of phospholipid were dried under nitrogen current. Dry lipid aliquots were then dissolved in 1 mL of n-hexane containing C19:0 as internal standard (0.75 μg mL−1, CAS number 1731-94-8, Merck, Darmstadt, Germany). Then, 200 μL of a methanolic solution of potassium hydroxide (2 mol L−1) were added to the tubes. The mixture was vortexed for 2 min and 2 mL of a saturated solution of sodium chloride were then added to each sample, followed by 5 min centrifugation at 626 × g. The upper was transferred into a microtube and dried under nitrogen current. Dried FAMEs were dissolved in 50 μL n-hexane and 2 μL of the resulting suspension were injected in an Agilent Technologies 6890 N Network Chromatograph (Santa Clara, CA, USA) equipped with a DB-FFAP column with 30 m length, an internal diameter of 0.32 mm and a film thickness of 0.25 μm (J & W Scientific, Folsom, CA, USA). Mass spectrometry analysis was performed in a connected Agilent 5973 Network Mass Selective Detector, operating at a 70 eV electron impact mode and scanning masses in the 50–550 m/z range, in a 1 s cycle and in full scan mode acquisition. The oven temperature was set at an initial temperature of 80 °C for 3 min, followed by three linear temperature increments to 160 °C at 25 °C min−1, to 210 °C at 2 °C min−1, and to 250 °C at 30 °C min−1. The temperature was finally set at 250 °C for 10 min. The injector and detector temperatures were respectively 220 and 280 °C. Helium was the carrier gas, at a flow rate of 1.3 mL min−1.49 Fatty acid identification was confirmed by comparing the experimental retention times and mass spectra to those obtained for the commercial FAME standards in the Supelco 37 Component FAME Mix (ref. 47885-U, Sigma-Aldrich, Darmstadt, Germany). The relative amount of each fatty acid was calculated as a relative percentage of the total area of the peaks, using the internal standard (C19:0) for normalization.Hydrophilic interaction liquid chromatography-mass spectrometry (HILIC-LC-MS)
Lipid extracts were analysed on high-performance liquid chromatographer (HPLC) Ultimate 3000 Dionex (Thermo Fisher Scientific, Bremen, Germany) coupled online to a Q-Exactive hybrid quadrupole mass spectrometer (Thermo Fisher, Scientific, Bremen, Germany). The elution protocol involved two mobile phases. The first mobile phase (A) consisted of water, acetonitrile, and methanol (25%, 50%, 25%) with 5 mM ammonium acetate, and the second (B) consisted of a mixture of acetonitrile and methanol (60%, 40%), with 5 mM ammonium acetate. Elution started with 5% of mobile phase A, held isocratically for 2 min followed by a linear increase to 48% within 8 min and then to 65% in 5 min and maintained for 2 min. Conditions were set to return to the initial settings in 13 min (3 min to decrease to 5% of phase A and a re-equilibration period of 10 min before the next injection). Elution into the Ascentis Si column HPLC Pore column (10 cm × 2.1 mm, 2.7 μm, Sigma-Aldrich) was carried out at a flow rate of 200 μL min−1, at 35 °C. For extract analysis, a volume of 5 μL of a mixture containing 10 μg of lipid extract (10 μL), 8 μL of phospholipid standards (0.04 μg dimyristoylphosphatidylcholine, 0.04 μg dimyristoylphosphatidylethanolamine, C19 lysophosphatidylcholine 0.04 μg, 0.08 μg dipalmitoylphosphatidylinositol, 0.024 μg dimyristoylphosphatidylglycerol, 0.08 μg dimyristoylphosphatidylserine, 0.16 μg tetramyristoylcardiolipin, 0.04 μg sphingomyelin (SM 17:0/d18:1), 0.16 μg dimyristoylphosphatidic acid, 0.08 μg of the ceramide Cer d35:1) and 82 μL of starting eluent were injected into the Ascentis Si column HPLC Pore column (10 cm × 2.1 mm, 2.7 μm, Sigma-Aldrich). Acquisitions in the Orbitrap® mass spectrometer were performed in both positive (electrospray voltage 3.0 kV) and negative (electrospray voltage −2.7 kV) modes, with a high resolution with 70![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 and AGC target of 1e6. The capillary temperature was 350 °C and the sheath gas flow was 20 U. For MS/MS determinations, the resolution was set to 17 500 and AGC target was set as 1e5 the cycles consisted of one full scan mass spectrum, and ten data-dependent MS/MS scans, with a dynamic exclusion of 60 seconds and intensity threshold of 2e4. Normalized collision energy™ (CE) was set to range between 25, 30 and 35 eV. Data acquisition was performed using the Xcalibur data system (V3.3, Thermo Fisher Scientific, USA). Phospholipid peak integration and assignments were performed using MZmine (version 2.32) using an in-house lipid species database. Only ions within 5 ppm were considered as correctly assigned. Polar lipid species were further confirmed by manual analyses of the MS/MS available spectra (Table S1, ESI†). Analyses of the MS/MS spectra acquired in the positive ion mode were performed to confirm the identity of the molecular species belonging to the Cer (ceramide), LPC (lysophosphatidylcholine), PC (phosphatidylcholine), LPE (lysophosphatidylethanolamine), PE (phosphatidylethanolamine) and SM (sphingomyelin) classes. The MS/MS spectra acquired in the negative ion mode were used to confirm the identity of LPC, PC, LPE, PE, LPG (lysophosphatidylglycerol), PG (phosphatidylglycerol), LPI (lysophosphatidylinositol), PI (phosphatidylinositol), PS (phosphatidylserine) and SM. Negative ion mode MS/MS data were used to identify the FA carboxylate anions fragments RCOO−, which allowed the assignment of the fatty acyl chains esterified to phospholipid precursors. The way this manual lipid species confirmation is performed, by detecting characteristic fragments in MS/MS spectra, has been described before for all phospholipid classes present in T. truncatus plasma.49 Relative percentages of lipid species per lipid class were calculated by exporting integrated peak areas values and upon normalization of the data by dividing the peak areas of the extracted ion chromatograms (XICs) of the polar lipid precursors of each class (listed in Table S1, ESI†) by the peak area of the internal standard selected for the class.
000 and AGC target of 1e6. The capillary temperature was 350 °C and the sheath gas flow was 20 U. For MS/MS determinations, the resolution was set to 17 500 and AGC target was set as 1e5 the cycles consisted of one full scan mass spectrum, and ten data-dependent MS/MS scans, with a dynamic exclusion of 60 seconds and intensity threshold of 2e4. Normalized collision energy™ (CE) was set to range between 25, 30 and 35 eV. Data acquisition was performed using the Xcalibur data system (V3.3, Thermo Fisher Scientific, USA). Phospholipid peak integration and assignments were performed using MZmine (version 2.32) using an in-house lipid species database. Only ions within 5 ppm were considered as correctly assigned. Polar lipid species were further confirmed by manual analyses of the MS/MS available spectra (Table S1, ESI†). Analyses of the MS/MS spectra acquired in the positive ion mode were performed to confirm the identity of the molecular species belonging to the Cer (ceramide), LPC (lysophosphatidylcholine), PC (phosphatidylcholine), LPE (lysophosphatidylethanolamine), PE (phosphatidylethanolamine) and SM (sphingomyelin) classes. The MS/MS spectra acquired in the negative ion mode were used to confirm the identity of LPC, PC, LPE, PE, LPG (lysophosphatidylglycerol), PG (phosphatidylglycerol), LPI (lysophosphatidylinositol), PI (phosphatidylinositol), PS (phosphatidylserine) and SM. Negative ion mode MS/MS data were used to identify the FA carboxylate anions fragments RCOO−, which allowed the assignment of the fatty acyl chains esterified to phospholipid precursors. The way this manual lipid species confirmation is performed, by detecting characteristic fragments in MS/MS spectra, has been described before for all phospholipid classes present in T. truncatus plasma.49 Relative percentages of lipid species per lipid class were calculated by exporting integrated peak areas values and upon normalization of the data by dividing the peak areas of the extracted ion chromatograms (XICs) of the polar lipid precursors of each class (listed in Table S1, ESI†) by the peak area of the internal standard selected for the class.
Data analysis
Phospholipid content and fatty acid results were presented as mean% ± standard deviation (STD). Respective graphs were produced using GraphPad Prism version 7.00 for Windows (GraphPad Software, La Jolla, California, USA). LC-MS data were glog transformed, EigenMS normalized,57 using the software R version 4.0.2.58 Principal Component Analysis (PCA) was conducted for exploratory data analysis, with the R package pcaMethods.59 Hierarchical clustering heatmaps were created using the R package pheatmap60 using “Euclidean” as clustering distance, and “ward.D” as the clustering method. Kruskal-Wallis test followed by Dunn's post hoc comparisons were performed with the R built-in function. p-Values were corrected for multiple testing using the BH Benjamini, Hochberg, and Yekutieli method (q values).61 Other R packages used for data management and graphics included ggplot2,62 dplyr,63 and tidyr64 (Fig. 1). | ||
| Fig. 1 Phospholipid content in T. truncatus in the polar lipids extracts from plasma (μg mL−1 of plasma). FI: female immature (N = 3); FM: female mature (N = 6); MI: male immature (N = 6); MM: male mature (N = 8). | ||
Results
Phospholipid quantification
Total phospholipid content in the polar lipid extract from plasma of T. truncatus were rather variable, ranging from 54.7 to 761.5 μg mL−1, with no statistically significant differences between groups. The values range from 191.7 ± 106.2 μg mL−1 in the female mature animals to 344.6 ± 105.6 μg mL−1 in the male immature group.Fatty acid profile characterization
The analysis of the T. truncatus fatty acid profile allowed the detection of 26 different fatty acids (Fig. 2). The more abundant fatty acids in the samples were palmitic acid (C16:0), stearic acid (C18:0) and oleic acid (C18:1n-9) in somewhat comparable amounts. The more abundant polyunsaturated fatty acids present were eicosapentaenoic acid (C20:5n-3) and docosahexaenoic acid (22:6n-3). Of notice is also the presence of odd-chain and branched FAs although in very small amounts (<1.5% for the more abundant C17:1n-7). There is a very obvious discrepancy between the fatty acid profiles of female and male mature animals. Of the 26 fatty acids present in the plasma of T. truncatus, the abundance of more than half (14) were statistically significant differences between these groups, including the aforementioned most prevalent species (C16:0, C18:0 and C18:1n-9). Importantly, the contents in several n-3 fatty acids, namely (C20:5n-3), docosapentaenoic acid (C22:5n-3) and (22:6n-3), were increased in mature males in comparison to mature females, as was the content in the n-6 fatty arachidonic acid (C20:4n-6). There were no statistically significant differences between immature specimens (female vs. male). Mature and immature female specimens differed between themselves in the fact that mature specimens had higher content in C16:0. The profiles of male specimens only differed in the content of C18:0, lower in adults (Fig. 2). | ||
| Fig. 2 Relative percentages (mean % ± standard 1 relative percentages (mean % ± standard deviation, STD) of the fatty acids present in T. truncatus polar lipid extracts from plasma samples. FI: female immature (N = 3); FM: female mature (N = 6); MI: male immature (N = 6); MM: male mature (N = 8). *p < 0.05; **p < 0.01; ***p < 0.001. | ||
Polar lipidome characterization
A lipidomics approach based on HILIC LC-MS allowed the identification and quantification of a total of 320 different lipid species (320 m/z ions) in the polar lipid extracts obtained from the plasma samples from T. truncatus specimens. There was a core of 314 lipid species that were present in the phospholipidome of all groups. A complete list of all lipid species detected in the study with information on their occurrence in each experimental group is available in Table S1 (ESI†).The detected species belonged to the following lipid classes: PC (105 lipid species), LPC (42), PE (67), LPE (18), PG (14), LPG (8), PI (14), LPI (2), PS (3), and SM (45) classes. Besides phospholipids, we were able to detect 2 Cer, namely Cer(d34:1) and Cer(d36:1). It is also relevant to highlight the number of alkylacyl phospholipids identified, mainly belonging to the PC (31 out of 105 species in total) and PE (35 out of 67) classes. The most abundant PC was PC(34:1), while the most abundant LPC was LPC(18:1) (Table S2, ESI†). Regarding the PE and LPE classes, the most abundant species in each were PE(38:5) and PE(38:6) and LPE(18:1). Unlike the case of the PC class, in which alkylacyl phospholipid forms were residual, in the PE class, there were several abundant alkylacyl PE lipid species, namely PE(O-38:6), PE(O-38:7), PE(O-40:6) and PE(O-40:7). For SM, the most abundant species in T. truncatus plasma was consistently SM(d34:1), while in the case of the remaining classes (PG, PI and PS) their abundance was more evenly distributed among the lipid species present in each class (see Table S2, ESI† for the complete relative quantitation results).
Principal component analysis (PCA) of the semi-quantification of the 320 different lipid species identified in this study resulted in a two-dimensional score plot accounting for 44.7% of the total variance, including the principal component 1 (27.6%) and principal component 2 (17.1%) (Fig. 3). The PCA plot shows discrimination between sexes, as well as the developmental stage, in the case of male subjects. A hierarchical clustering heat map analysis including the 50 most significant polar lipid discriminating species after Kruskal-Wallis (KW) statistical analysis (including the 50 phospholipids with the lowest p values) allowed graphically visualizing lipid species particularly contributing to the discrimination between groups (Fig. 4). The heatmap is divided in four different zones highlighting differences between groups. The two lower leaflets of the heatmap allow to observe a clear separation between sexes driven mainly mainly the content of PE alkylacyl lipids, but also of certain species of LPC and LPE, which were increased in males (bottom right leaflet of the heat map). In the upper right leaflet of the heatmap, dissimilarities between immature and mature male individuals become evident, driven in particular by the contents of PC species with a total number of 31, 32, and 33 carbons in their acyl chains, but also the contents of three SM species, all increased in the mature male group. Finally, discriminating between the immature and mature female groups is not as obvious, but it is possible to observe an increased content in all PS in immature females compared to mature females (upper left leaflet of the heatmap), as well as an increased content in a group of PE alkylacyl lipids (lower part of the lower left leaflet of the heatmap). Fig. 5 shows the box plots of the 16 most significant different lipid species (Kruskal-Wallis, q < 0.01). Among these 16 lipid species with lowest q-value, we have a clear predominance of PE alkylacyl phospholipids (9 out of the sixteen), as well as lysophospholipids (2 LPE, 1 LPC) and two of the three PS species detected.
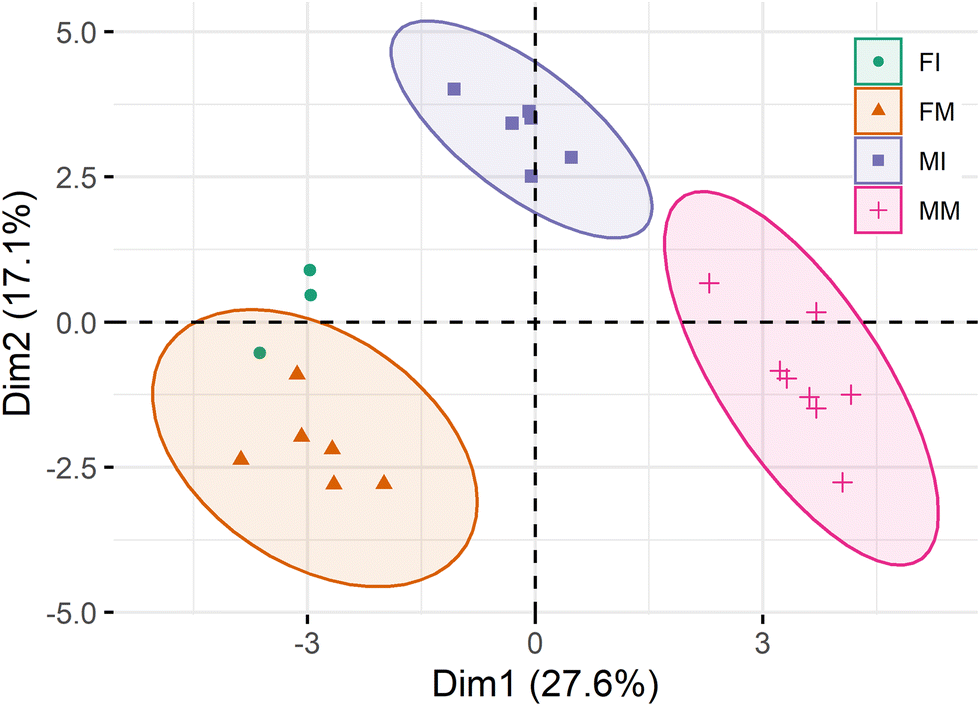 | ||
| Fig. 3 Principal component analysis (PCA) of the relative quantitative analysis of the phospholipidome of T. truncatus subdivided according to sex and developmental stage. FI: female immature (N = 3); FM: female mature (N = 6); MI: male immature (N = 6); MM: male mature (N = 8). | ||
 | ||
| Fig. 4 Two-dimensional hierarchical clustering heat map with dendrogram, of the 50 most significant polar lipid discriminating species after Kruskal-Wallis (KW) statistical analysis. Normalized relative abundance levels are shown on the color scale, with numbers indicating the fold difference from the mean. FI: female immature; FM: female mature; MI: male immature; MM: male mature. | ||
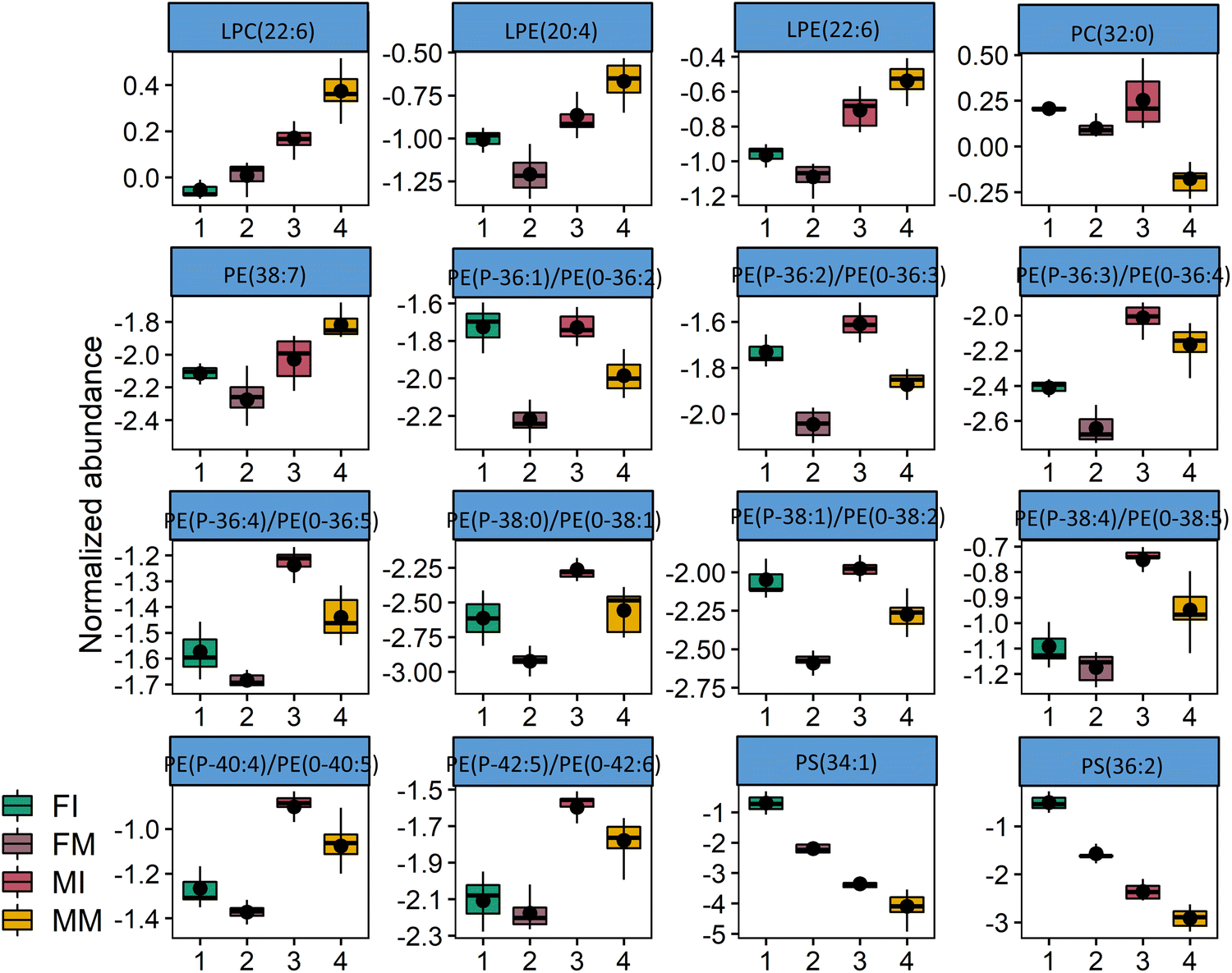 | ||
| Fig. 5 Box plots of the 16 most significant different lipid species (Kruskal-Wallis, p < 0.01). FI: female immature (N = 3); FM: female mature (N = 6); MI: male immature (N = 6); MM: male mature (N = 8). | ||
In Table 2, all the lipid species that were found to be statistically different between the groups (Kruskal-Wallis test, followed by Dunn's multiple comparison test, with FDR correction) are comprehensively listed. It is possible to observe that most of the phospholipids that are statistically different belong to the LPC and LPE classes (a total of 9), and four different PE species. When comparing immature and mature males, the class containing more statistically significant differences is PC (10 statistically different species), followed by SM (4). When analysing statistically different lipid species between the immature male and female groups, the most represented lipid class was PE, more specifically PE alkylacyl lipids (6 statistically different species) followed by LPE (3). Finally, between immature and mature females, the lipid class displaying the most statistically different species was also PE (7), and again especially PE alkylacyl lipids (4). These observations mainly confirm the classes that contributed the most for group discrimination in the two-dimensional hierarchical clustering heat map (Fig. 4).
| FI vs. FM | p | p.adj | FI vs. MI | p | p.adj |
|---|---|---|---|---|---|
| FI: female immature (N = 3); FM: female mature (N = 6); MI: male immature (N = 6); MM: male mature (N = 8). | |||||
| PE(P-40:2)/PE(O-40:3) | 0.000581 | 0.012118 | LPE(O-16:0) | 0.001229 | 0.01784 |
| SM(d30:0) | 0.001229 | 0.01784 | PC(P-28:0)/PC(O-28:1) | 0.002499 | 0.026494 |
| PE(32:1) | 0.001388 | 0.019078 | PC(O-28:0) | 0.002802 | 0.028311 |
| PG(34:4) | 0.001564 | 0.020433 | PE(P-42:5)/PE(O-42:6) | 0.003921 | 0.03422 |
| PE(P-36:1)/PE(O-36:2) | 0.001762 | 0.021544 | LPC(O-16:0) | 0.004879 | 0.038457 |
| SM(d30:1) | 0.002802 | 0.028311 | LPE(O-18:0) | 0.005433 | 0.040905 |
| PE(36:2) | 0.003137 | 0.029968 | PE(P-38:5)/PE(O-38:6) | 0.005433 | 0.040905 |
| PE(P-38:2)/PE(O-38:3) | 0.003137 | 0.029968 | LPC(O-14:0) | 0.006714 | 0.045692 |
| PG(32:1) | 0.003509 | 0.032393 | LPC(P-16:0)/LPC(O-16:1) | 0.006714 | 0.045692 |
| PE(P-38:1)/PE(O-38:2) | 0.004879 | 0.038457 | LPE(P-16:0)/LPE(O-16:1) | 0.006714 | 0.045692 |
| PE(32:2) | 0.007452 | 0.048667 | PE(P-36:5)/PE(O-36:6) | 0.006714 | 0.045692 |
| PE(P-40:5)/PE(O-40:6) | 0.006714 | 0.045692 | |||
| PE(P-38:4)/PE(O-38:5) | 0.007452 | 0.048667 | |||
| MI vs. MM | p | p.adj | FM vs. MM | p | p.adj |
|---|---|---|---|---|---|
| SM(d38:3) | 4.43 × 10−5 | 0.006637 | PE(38:7) | 3.46 × 10−5 | 0.006637 |
| SM(d37:2) | 5.38 × 10−5 | 0.006637 | LPE(20:4) | 3.82 × 10−5 | 0.006637 |
| PI(36:4) | 5.93 × 10−5 | 0.006637 | LPE(22:6) | 4.02 × 10−5 | 0.006637 |
| SM(d38:2) | 7.91 × 10−5 | 0.006637 | LPE(22:5) | 7.54 × 10−5 | 0.006637 |
| PE(39:6) | 8.29 × 10−5 | 0.006637 | PC(P-42:7)/PC(O-42:8) | 7.91 × 10−5 | 0.006637 |
| PC(33:0) | 9.55 × 10−5 | 0.006637 | PE(40:7) | 0.00011 | 0.006637 |
| PC(P:34:0)/PC(O-34:1) | 0.000121 | 0.006637 | LPC(20:4) | 0.000166 | 0.00726 |
| PC(32:0) | 0.000132 | 0.006637 | LPE(20:5) | 0.000166 | 0.00726 |
| PC(40:3) | 0.000132 | 0.006637 | LPC(22:6) | 0.000218 | 0.007903 |
| PC(P-36:1)/PC(O-36:2) | 0.000132 | 0.006637 | PE(40:6) | 0.000218 | 0.007903 |
| SM(d40:3) | 0.000132 | 0.006637 | LPC(22.5) | 0.000239 | 0.008179 |
| PC(30:0) | 0.000182 | 0.007443 | SM(d33:0) | 0.000239 | 0.008179 |
| PC(O-34:0) | 0.000182 | 0.007443 | LPE(16.0) | 0.000249 | 0.008336 |
| PC(P-32:0)/PC(O-32:1) | 0.000199 | 0.007903 | PE(40:5) | 0.000261 | 0.008484 |
| PC(P-32:1)/PC(O-32:2) | 0.000209 | 0.007903 | PG(34:2) | 0.000354 | 0.009514 |
| PC(31:0) | 0.000273 | 0.008577 | LPE(18:1) | 0.00037 | 0.009514 |
Discussion
Plasma levels of lipids, lipoproteins and particular molecular components, including polar lipids are very tightly regulated in mammals and kept within specific ranges in health conditions.65 Variation in lipoproteins levels have been correlated with disease in dolphins and proved useful to detect unhealthy conditions in these animals,21 but while the variation in plasma/serum lipids has been well studied in humans, it was so far unexplored in cetaceans. Polar lipids circulate in the plasma as components of lipoproteins,66 accounting for approximately 27% of the total lipid content of T. truncatus and including mainly phospholipids.67,68 Our determinations of the plasma phospholipid concentration values of T. truncatus, although highly variable even within experimental groups, were generally comparable to values reported in humans and other terrestrial mammal species.69–71 This variation may be the result of individual characteristics and inter-individual metabolic differences, but may also be linked to differences in diet, or even to different periods after the last meal before sampling.This is the first work looking into how cetacean plasma polar lipid content is modulated by sex and developmental stage. Several studies have proposed general metabolic changes in T. truncatus metabolism taking place with sex and age39,40,45–48 but this is the first insight into plasma lipid regulation by individual features. Each experimental group (female immature, female adults, male immature and male adults) was shown to display underlined characteristics that made them unique and easily distinguishable from the others in terms of fatty acid and polar lipid compositions (Fig. 6). In terms of fatty acid profiles, some similarities take place, namely the occurrence of the same main fatty acids (palmitic (C16:0), stearic (C18:0) and oleic (C18:1n-9) acids) throughout groups. However, the main observation here are the marked differences between adults from different sexes, with more than half of the detected fatty acid species occurring in statistically different amounts. The lipidomics analysis of the plasma polar lipid content of the dolphins highlighted marked differences not only between adults from different sexes, but throughout all groups, making it possible to clearly differentiate/discriminate between them considering the occurrence of specific lipid species. The results regarding the fatty acid profiles in this study are generally in line with what we had described in a previous study characterizing the plasma of T. truncatus, both in terms of species represented and most abundant species.49 Eicosapentaenoic (C20:5n-3) and docosahexaenoic (C22:6n-3) acids are present at significantly higher amounts in dolphins, while some n-6 fatty acids, particularly arachidonic acid (20:4n-6), are more abundant in terrestrial mammals.31,72 This fact, together with the presence of particular fatty acids, namely the fatty acids with odd or branched carbon chains are in the plasma of T. truncatus but not present in their terrestrial counterparts, has been suggested as evidence that these cetaceans may absorb and utilize lipids from fish, or have a more complex lipid metabolism.20,24,25
 | ||
| Fig. 6 Main differences in fatty acid profile and phospholipid composition in the plasma of T. truncatus specimens grouped by sex and developmental stage. | ||
The lipidomics analysis of the polar fraction of T. truncatus plasma samples allowed the identification and relative quantification of a total of 320 different lipid species (320 m/z ions). These results generally confirmed our previous observations in T. truncatus plasma in terms of major species in the most represented phospholipid classes, as well as a clear divergence from terrestrial mammals in terms of polar lipid composition.49 The most abundant PC species in T. truncatus plasma was PC(34:1) in all experimental groups, which is consistent with our previous observations in adult males.49 The most abundant PC alkylacyl phospholipid species was PC(P-38:5)/PC(O-38:6) in all groups, also confirming our previous results, but PC alkylacyl occur in much lower abundance when compared with the major PC species. The most abundant LPC species is LPC(18:1) in all groups, again in agreement with our previous report.49 Regarding the PE class, the most abundant species were PE(38:6) followed by PE(38:5) in all groups, while PE alkylacyl were also detected, representing a significant part of the total content of the class. The main PE alkylacyl present were PE(P-38:5)/PE(O-38:6), PE(P-38:6)/PE(O-38:7) and PE(P-40:6)/PE(O-40:7), present in amounts comparable to the most abundant non-ether lipids of the PE class.49 The most abundant LPE in T. truncatus plasma was confirmed to be LPE(18:1), in all experimental groups. Regarding the SM class, the main class of sphingolipids in T. truncatus plasma, the most abundant species were SM(d34:1) followed by SM(d36:1) in all groups, has also reported previously in adult males.49
Male adults differ profoundly from female adults in terms of fatty acid profiles. Those differences include the most abundant species (C16:0, C18:0 and C18:1n-9), all in higher contents in the mature female groups. Given its importance as a prime constituent of triacylglycerols in adipose tissue in humans, palmitic acid has been proposed as having the greatest diagnostic and prognostic significance, while also being linked to adverse cardiovascular events.73–75 Since, adipose deposits (blubber specifically), and therefore body composition have been reported to be heavily influenced sex (and age) in bottlenose dolphins,76 palmitic acid content may be a particularly interesting parameter in to consider in these dolphins. Unlike palmitic acid, stearic does not seem to have a significant influence on lipid metabolism and is not considered a primary risk factor for cardiovascular disease.94–98 As for oleic acid it has also been ascribed generally beneficial effects, including improving the blood lipid profile,99 and contributing for the maintenance of a healthy body weight and prevent metabolic syndrome manifestations.100 Adult male specimens also present increased content in several n-3 fatty acids, including eicosapentaenoic and docosapentaenoic acids, while displaying lower content of n-6 arachidonic acid. While the first two have been consistently ascribed beneficial effects namely in the contexts of hypertension,77 hyperlipidemia,78 hyperglycemia,79 insulin sensitivity,80 and metabolic syndrome manifestations in general,81 arachidonic acid has been attributed deleterious effects, namely the promotion, development and progression of cardiovascular diseases and cancer.82 Adult males also displayed characteristics in phospholipid plasma content that made them differ from other groups. In this case, adult male specimens displayed the lowest contents of PC species with total number of 31, 32, and 33 carbons in their acyl chains, but also the contents of three SM species. Two of the SM species of the top 50 discriminating species contained odd acyl chains (SM(d33:0) and SM(d37:2)). Plasma SM content has been reported to increase with a high-fat diet feeding.86–89 In humans, SM levels have been proposed to be an important predictor or an early sign of pre-diabetes and a biomarker for the development of diabetes.90 Plasma SM has also been proposed as an independent risk factor for coronary heart disease.91,92 Moreover, adult males also displayed the highest amounts in some lysolipids species, particularly those carrying essential fatty acids ((LPC(20:4), LPC(22:5), LPC(22:6), LPC(24:5), LPC(24:6), LPE(20:4), LPE(22:5) and LPE(22:6)). Plasma LPC has been reported to have inflammatory, anti-hemostatic, cytotoxic and atherogenesis promoting roles.83 Some of the previously mentioned LPC species, namely polyunsaturated ones and those containing docosahexaenoic acid, have been attributed to anti-inflammatory activity.84,85 As for LPE species, those containing docosahexaenoic acid, in particular, are very abundant in the brain,83 therefore their plasma content may be physiologically important.
Mature females also displayed idiosyncratic features in terms of polar lipid species content, namely displaying the lowest amounts of in certain LPC and LPE (most abundant in mature males), as well as the lowest abundance in many PE alkylacyl lipids. PE ether lipids were proposed roles as scavengers of free radicals and singlet oxygen.101,102 Moreover, PE plasmalogens containing n-3 PUFA have been proposed as showing potential in the prevention of neurodegenerative diseases, namely Alzheimer's disease,103 Parkinson's disease,104 while also alleviating atherosclerosis processes.105 Therefore, these differences driven by sex in T. truncatus may have very relevant physiological meaning, mainly in aging individuals.
In contrast with the mature female group, immature males group display the higher amounts in the same PE alkylacyl lipids and differ greatly from mature males in terms of PC species with total number of 31, 32, and 33 carbons in their acyl chains. Incorporation of odd chain or branched fatty acids in plasma lipids was suggested as a strong sign of uptake of these fatty acids from fish and of a more complex lipid metabolism in these dolphins when compared to terrestrial counterparts.106 Therefore, the observed changes may reflect changes in lipid metabolism with age, which would be especially evident in males.
Finally, the main differentiating characteristic of the female immature group is a remarkably increased content in the three PS species detected (PS(34:1), PS(34:2) and PS(36:2)) regarding other groups. This may have physiological significance since PS has been shown to be selectively enriched in small high-density lipoprotein (HDL) vesicles, actively contributing to HDL-mediated atheroprotection,93 although this should not be particularly important in younger individuals.
Studies have reported that, in T. truncatus, energy intake and total energy expenditure decrease with age, while fat mass increases.39,40 Other studies have reported the influence of age and sex on blood, serum, and plasma biochemical variables in T. truncatus.45–47 Another recent study involving an impressive number of specimens and samples confirmed this influence and identified numerous effects of age and sex on the concentration of several analytes in the plasma of T. truncatus.48 This study, comprising 1426 blood samples collected from 156 different dolphins, measured 37 analytes in each sample. Mean platelet volume, glucose, blood urea nitrogen, globulins, gamma-glutamyl transpeptidase activity and chloride, which increase with age, while absolute lymphocytes, total protein, alkaline phosphatase activity, creatinine kinase activity, and calcium, all decreased with age. Moreover, a number of analytes/parameters were found to be influenced by sex, with females displaying higher median iron, absolute lymphocytes and mean corpuscular haemoglobin concentration. The physiological differences with age and sex observed in plasma analytes were also observed in this work regarding lipid composition. However, to assess the significance of the differences described here as elicited by sex and age, more efforts to study and clarify the particular lipid metabolism of cetaceans must be planned and expedited. Regarding the limitations of this work, we have to assume that animal diet may have an effect in the results. The influence of diet in plasma lipid composition has been well documented in these animals, and fatty acid composition has been shown to be especially susceptible.19,20,107 The fact is that, despite the daily fish offer being generally the same for all the specimens in the park, individuals may display specific preferences between the fish species made available what might have contributed to individual variability. Moreover, diet may be a factor in establishing the differences between immature and mature individuals especially in the youngest specimens, since there is a period of dietetic transition (a mixt diet of milk and fish), as the several fish species are introduced gradually in the diet. It would be very interesting to study how diet (specific fish species, for instance) and fasting (even if intermittent) reverberate in the plasma lipid composition and even in endocrine signaling. This would certainly provide important new clues about lipid metabolism regulation and lipid incorporation through diet, while also allowing a better understanding of these phenomena as occurring in the wild, for instance during cetacean migration. Another enlightening approach could be the study of how extensively and quickly plasma lipid content changes in rehabilitating wild animals as subjected to a controlled homogenized diet. Nevertheless, given the ethical considerations and even technical constraints, the study of managed cetaceans still represents the most practical way to get meaningful insight into cetacean metabolism.
Conclusions
This study contributes to further understand the physiology of small cetaceans by unveiling developmental stage and sex modulation of the plasma phospholipidome. The data presented here may provide a useful interpretation baseline for lipid analytes for upcoming comparative studies, either involving managed delphinids of different sexes and ages.Author contributions
Conceptualization, J. P. M. and M. R. D.; methodology, investigation and resources, J. P. M., H. B. F., T. M., C. F., N. U., J. N. and M. R. D.; writing – original draft preparation, J. P. M. and M. R. D.; writing – review & editing, P. D.; all authors reviewed the manuscript.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This is a contribution of the Marine Lipidomics Laboratory, University of Aveiro. The authors are grateful to FCT/MCTES (Portugal) for the financial support to CESAM (UIDB/50017/2020 + UIDP/50017/2020 + LA/P/0094/2020), LAQV/REQUIMTE (UIDB/50006/2020) and RNEM (LISBOA-01-0145-FEDER-402-022125), through national funds, and the co-funding by the FEDER, within the PT2020 Partnership Agreement and Compete 2020. The authors would like to thank Zoomarine Portugal for providing the samples and the training and veterinary staff for the time they spent to make this study possible. Helena B. Ferreira is grateful to FCT for her PhD Grant (2020.04611.BD). Tânia Melo thanks the junior research contract in the scope of the Individual Call to Scientific Employment Stimulus 2020 (CEECIND/01578/2020).References
- A. Serres, Y. Hao and D. Wang, Behav. Processes, 2020, 170, 103998 CrossRef PubMed.
- R. S. Wells and M. D. Scott, in Encyclopedia of Marine Mammals, ed. B. Würsig, J. G. M. Thewissen and K. M. Kovacs, Academic Press, 3rd edn, 2018, pp. 118–125 DOI:10.1016/B978-0-12-804327-1.00072-8.
- N. B. Barros and D. K. Odell, in The Bottlenose Dolphin, ed. S. L. R. R. Reeves, Academic Press, San Diego, CA, 1990, pp. 309–328 Search PubMed.
- J. G. Mead and C. W. Potter, in The Bottlenose Dolphin, ed. S. L. R. R. Reeves, Academic Press, San Diego, CA., 1990, pp. 165–195 Search PubMed.
- T. Segawa, H. Amatsuji, K. Suzuki, M. Suzuki, M. Yanagisawa, T. Itou, T. Sakai and T. Nakanishi, Results Immunol., 2013, 3, 57–63 CrossRef PubMed.
- K. E. Hunt, M. J. Moore, R. M. Rolland, N. M. Kellar, A. J. Hall, J. Kershaw, S. A. Raverty, C. E. Davis, L. C. Yeates, D. A. Fauquier, T. K. Rowles and S. D. Kraus, Conserv. Physiol., 2013, 1, cot006 Search PubMed.
- F. Terasawa, M. Kitamura, A. Fujimoto and S.-I. Hayama, J. Vet. Med. Sci., 2002, 64, 1075–1078 CrossRef PubMed.
- T. B. S. Group, Circulation, 2000, 102, 21–27 CrossRef PubMed.
- O. Quehenberger and E. A. Dennis, N. Engl. J. Med., 2011, 365, 1812–1823 CrossRef CAS PubMed.
- S. Rai and S. Bhatnagar, OMICS: J. Integr. Biol., 2017, 21, 132–142 CrossRef CAS PubMed.
- A. Sigruener, M. E. Kleber, S. Heimerl, G. Liebisch, G. Schmitz and W. Maerz, PLoS One, 2014, 9, e85724 CrossRef PubMed.
- H. Hinterwirth, C. Stegemann and M. Mayr, Circ.: Cardiovasc. Genet., 2014, 7, 941–954 CAS.
- Z. H. Alshehry, P. A. Mundra, C. K. Barlow, N. A. Mellett, G. Wong, M. J. McConville, J. Simes, A. M. Tonkin, D. R. Sullivan, E. H. Barnes, P. J. Nestel, B. A. Kingwell, M. Marre, B. Neal, N. R. Poulter, A. Rodgers, B. Williams, S. Zoungas, G. S. Hillis, J. Chalmers, M. Woodward and P. J. Meikle, Circulation, 2016, 134, 1637–1650 CrossRef CAS PubMed.
- S. Kohno, A. L. Keenan, J. M. Ntambi and M. Miyazaki, Biochem. Biophys. Res. Commun., 2018, 504, 590–595 CrossRef CAS PubMed.
- Y. K. Tham, B. C. Bernardo, K. Huynh, J. Y. Y. Ooi, X. M. Gao, H. Kiriazis, C. Giles, P. J. Meikle and J. R. McMullen, Cell Rep., 2018, 24, 2757–2772 CrossRef CAS PubMed.
- H. Huang, G. Ye, S.-Q. Lai, H.-X. Zou, B. Yuan, Q.-C. Wu, L. Wan, Q. Wang, X.-L. Zhou, W.-J. Wang, Y.-P. Cao, J.-F. Huang, S.-L. Chen, B.-C. Yang and J.-C. Liu, Frontiers in Cardiovascular Medicine, 2021, 8, 757022 CrossRef CAS PubMed.
- T. Hornemann, in Prevention and Treatment of Atherosclerosis: Improving State-of-the-Art Management and Search for Novel Targets, ed. A. von Eckardstein and C. J. Binder, Springer International Publishing, Cham, 2022, pp. 493–510 DOI:10.1007/164_2021_517.
- P. M. Sobolesky, T. S. Harrell, C. Parry, S. Venn-Watson and M. G. Janech, Front. Endocrinol., 2016, 7, 33 Search PubMed.
- S. K. Venn-Watson, C. Parry, M. Baird, S. Stevenson, K. Carlin, R. Daniels, C. R. Smith, R. Jones, R. S. Wells, S. Ridgway and E. D. Jensen, PLoS One, 2015, 10, e0132117 CrossRef PubMed.
- M. Suzuki, M. Yoshioka, Y. Ohno and Y. Akune, Sci. Rep., 2018, 8, 12030 CrossRef PubMed.
- S. Venn-Watson, C. R. Smith, S. Stevenson, C. Parry, R. Daniels, E. Jensen, V. Cendejas, B. Balmer, M. Janech, B. A. Neely and R. Wells, Front. Endocrinol., 2013, 4, 136 CrossRef PubMed.
- D. S. Houser, D. Derous, A. Douglas and D. Lusseau, J. Exp. Biol., 2021, 224, jeb238915 CrossRef PubMed.
- Y. Endo, K.-i Kamei and M. Inoue-Murayama, J. Evol. Biol., 2018, 31, 1655–1665 CrossRef CAS PubMed.
- Z. Wang, Z. Chen, S. Xu, W. Ren, K. Zhou and G. Yang, Sci. Rep., 2015, 5, 14187 CrossRef PubMed.
- Z. Swaim, A. J. Westgate, H. N. Koopman, R. Rolland and S. Kraus, Endangered Species Research, 2009, 6, 259–271 CrossRef.
- K. M. Wynn, J. Jamison, R. Richter, S. E. Woods, R. Partha, A. Kowalczyk, C. Kronk, M. Chikina, R. K. Bonde, D. E. Crocker, J. Gaspard, J. M. Lanyon, J. Marsillach, C. E. Furlong and N. L. Clark, Science, 2018, 361, 591–594 CrossRef PubMed.
- D. Derous, J. Sahu, A. Douglas, D. Lusseau and M. Wenzel, Conserv. Physiol., 2021, 9, coaa136 CrossRef CAS PubMed.
- G. Di Guardo, Res. Vet. Sci., 2018, 119, 43–44 CrossRef PubMed.
- S. Venn-Watson, E. D. Jensen and N. J. Schork, Proc. Natl. Acad. Sci. U. S. A., 2020, 117, 20950–20958 CrossRef CAS PubMed.
- F. E. Scaglione, E. Bollo, P. Pregel, L. Chiappino, A. Sereno, W. Mignone, R. Moschi, F. Garibaldi, C. Tittarelli and F. Guarda, Dis. Aquat. Org., 2013, 107, 31–36 CrossRef CAS PubMed.
- W. S. Harris and T. L. Schmitt, Prostaglandins, Leukotrienes Essent. Fatty Acids, 2014, 90, 55–59 CrossRef CAS PubMed.
- L. M. Mazzaro, S. P. Johnson, P. A. Fair, G. Bossart, K. P. Carlin, E. D. Jensen, C. R. Smith, G. A. Andrews, P. S. Chavey and S. Venn-Watson, Comp. Med., 2012, 62, 508–515 CAS.
- S. Venn-Watson, C. Benham, K. Carlin, D. DeRienzo and J. St Leger, J. Zoo Wildl. Med., 2012, 43, S35–47 CrossRef PubMed.
- S. K. Venn-Watson and S. H. Ridgway, Comp. Med., 2007, 57, 390–395 CAS.
- S. Venn-Watson, Front. Endocrinol., 2014, 5, 227 Search PubMed.
- S. Venn-Watson, K. Carlin and S. Ridgway, Gen. Comp. Endocrinol., 2011, 170, 193–199 CrossRef CAS PubMed.
- D. Gunn-Moore, O. Kaidanovich-Beilin, M. C. Gallego Iradi, F. Gunn-Moore and S. Lovestone, Alzheimers Dementia, 2018, 14, 195–204 CrossRef PubMed.
- I. Stylianaki, A. Komnenou, D. Posantzis, K. Nikolaou and N. Papaioannou, Veterinary Record Case Reports, 2019, 7, e000700 CrossRef.
- R. Rimbach, A. Amireh, A. Allen, B. Hare, E. Guarino, C. Kaufman, H. Salomons and H. Pontzer, J. Exp. Biol., 2021, 224, jeb242218 CrossRef PubMed.
- M. Reddy, T. Kamolnick, C. Curry and D. Skaar, Marine Mammals: Public Display and Research, 1994, 1, 26–31 Search PubMed.
- P. A. Fair, E. Montie, L. Balthis, J. S. Reif and G. D. Bossart, Gen. Comp. Endocrinol., 2011, 174, 184–194 CrossRef CAS PubMed.
- C. A. Thomson and J. R. Geraci, Can. J. Fish. Aquat. Sci., 1986, 43, 1010–1016 CrossRef.
- D. J. St. Aubin, S. H. Ridgway, R. S. Wells and H. Rhinehart, Mar. Mammal Sci., 1996, 12, 1–13 CrossRef.
- M. Suzuki, S. Uchida, K. Ueda, T. Tobayama, E. Katsumata, M. Yoshioka and K. Aida, Gen. Comp. Endocrinol., 2003, 132, 427–433 CrossRef CAS PubMed.
- L. K. Lauderdale, M. T. Walsh, K. A. Mitchell, D. A. Granger, J. D. Mellen and L. J. Miller, PLoS One, 2021, 16, e0250332 CrossRef CAS PubMed.
- A. J. Hall, R. S. Wells, J. C. Sweeney, F. I. Townsend, B. C. Balmer, A. A. Hohn and H. L. Rhinehart, Comp. Biochem. Physiol., Part A: Mol. Integr. Physiol., 2007, 148, 266–277 CrossRef PubMed.
- J. D. Goldstein, E. Reese, J. S. Reif, R. A. Varela, S. D. McCulloch, R. H. Defran, P. A. Fair, G. D. Bossart and L. Hansen, J. Wildl. Dis., 2006, 42, 447–454 CrossRef PubMed.
- H. H. Nollens, N. J. Haney, N. I. Stacy and T. R. Robeck, Vet. Q., 2020, 40, 342–352 CrossRef CAS PubMed.
- J. P. Monteiro, E. Maciel, T. Melo, C. Flanagan, N. Urbani, J. Neves and M. R. Domingues, Lipids, 2021, 56, 461–473 CrossRef CAS PubMed.
- C.-H. Tang, C.-Y. Lin, Y.-L. Tsai, S.-H. Lee and W.-H. Wang, Zoo Biol., 2018, 37, 440–451 CrossRef CAS PubMed.
- I. M. Ruiz-Hernández, M.-Z. Nouri, M. Kozuch, N. D. Denslow, R. E. Díaz-Gamboa, R. Rodríguez-Canul and R. C. Collí-Dulá, Chemosphere, 2022, 299, 134353 CrossRef PubMed.
- P. Bories, A. H. Rikardsen, P. Leonards, A. T. Fisk, S. Tartu, E. F. Vogel, J. Bytingsvik and P. Blévin, Ecol. Evol., 2021, 11, 6716–6729 CrossRef PubMed.
- S. Brando, D. M. Broom, C. Acasuso-Rivero and F. Clark, Behav. Processes, 2018, 156, 16–36 CrossRef PubMed.
- E. M. Bartlett and D. H. Lewis, Anal. Biochem., 1970, 36, 159–167 CrossRef CAS PubMed.
- J. P. Monteiro, T. Melo, J. Skjermo, S. Forbord, O. J. Broch, P. Domingues, R. Calado and M. R. Domingues, Algal Res., 2021, 54, 102201 CrossRef.
- J. P. Monteiro, E. Maciel, R. Maia, A. T. Pereira, R. Calado, P. Domingues, T. Melo, C. Eira and M. R. Domingues, Mar. Mammal Sci., 2021, 37, 1406–1427 CrossRef CAS.
- Y. V. Karpievitch, S. B. Nikolic, R. Wilson, J. E. Sharman and L. M. Edwards, PLoS One, 2014, 9, e116221 CrossRef PubMed.
- R. C. Team, R: A language and environment for statistical computing, R Foundation for Statistical Computing, Vienna, Austria, 2013 Search PubMed.
- W. Stacklies, H. Redestig, M. Scholz, D. Walther and J. Selbig, Bioinformatics, 2007, 23, 1164–1167 CrossRef CAS PubMed.
- R. Kolde, R package version 1.0.12, 2019 Search PubMed.
- Y. Benjamini and Y. Hochberg, J. R. Stat. Soc. Series B Stat. Methodol., 1995, 57, 289–300 Search PubMed.
- H. Wickham, ggplot2: elegant graphics for data analysis, Springer-Verlag, New York, 2016 Search PubMed.
- H. Wickham, R. François, L. Henry and K. Müller, dplyr: A Grammar of Data Manipulation, R package version 0.7.7, 2018.
- H. Wickham and L. Henry, R package version 0.8.1, 2018, 3 Search PubMed.
- K. Kawasumi, Y. Hirakawa, P. Lee, N. Mori, I. Yamamoto, T. Arai and F. Terasawa, J. Anim. Vet. Adv., 2012, 11, 3781–3784 Search PubMed.
- S. Rauschert, A. Gázquez, O. Uhl, F. F. Kirchberg, H. Demmelmair, M. Ruíz-Palacios, M. T. Prieto-Sánchez, J. E. Blanco-Carnero, A. Nieto, E. Larqué and B. Koletzko, Lipids Health Dis., 2019, 18, 20 CrossRef PubMed.
- G. J. Nelson, Comparative Biochemistry and Physiology Part B: Comparative Biochemistry, 1971, 40, 423–432 CrossRef CAS PubMed.
- G. J. Nelson, Comparative Biochemistry and Physiology Part B: Comparative Biochemistry, 1973, 46, 257–268 CrossRef CAS PubMed.
- G. J. Nelson, Lipids, 1967, 2, 323–328 CrossRef CAS.
- O. Quehenberger, A. M. Armando, A. H. Brown, S. B. Milne, D. S. Myers, A. H. Merrill, S. Bandyopadhyay, K. N. Jones, S. Kelly, R. L. Shaner, C. M. Sullards, E. Wang, R. C. Murphy, R. M. Barkley, T. J. Leiker, C. R. H. Raetz, Z. Guan, G. M. Laird, D. A. Six, D. W. Russell, J. G. McDonald, S. Subramaniam, E. Fahy and E. A. Dennis, J. Lipid Res., 2010, 51, 3299–3305 CrossRef CAS PubMed.
- G. B. Phillips and J. T. Dodge, J. Lipid Res., 1967, 8, 676–681 CrossRef CAS.
- L. Hodson, C. M. Skeaff and B. A. Fielding, Prog. Lipid Res., 2008, 47, 348–380 CrossRef CAS PubMed.
- Y. Li, A. Hruby, A. M. Bernstein, S. H. Ley, D. D. Wang, S. E. Chiuve, L. Sampson, K. M. Rexrode, E. B. Rimm, W. C. Willett and F. B. Hu, J. Am. Coll. Cardiol., 2015, 66, 1538–1548 CrossRef CAS PubMed.
- C. L. Chei, K. Yamagishi, A. Kitamura, M. Kiyama, T. Sankai, T. Okada, H. Imano, T. Ohira, R. Cui, M. Umesawa, I. Muraki, T. Tanigawa, S. Sato and H. Iso, Circulation journal: official journal of the Japanese Circulation Society, 2018, 82, 3013–3020 CrossRef CAS PubMed.
- M. E. Kleber, G. E. Delgado, C. Dawczynski, S. Lorkowski, W. März and C. von Schacky, J. Clin. Lipidol., 2018, 12, 455–463.e453 CrossRef PubMed.
- S. K. Adamczak, R. R. Holser, D. P. Costa, E. J. B. McCabe and R. S. Wells, Frontiers in Marine Science, 2021, 8, 615773 CrossRef.
- C. I. Bercea, G. S. Cottrell, F. Tamagnini and A. J. McNeish, Br. J. Pharmacol., 2021, 178, 860–877 CrossRef CAS PubMed.
- A. C. Skulas-Ray, P. W. F. Wilson, W. S. Harris, E. A. Brinton, P. M. Kris-Etherton, C. K. Richter, T. A. Jacobson, M. B. Engler, M. Miller, J. G. Robinson, C. B. Blum, D. Rodriguez-Leyva, S. D. de Ferranti and F. K. Welty, Circulation, 2019, 140, e673–e691 CrossRef CAS PubMed.
- P. Zhuang, H. Li, W. Jia, Q. Shou, Y. E. Zhu, L. Mao, W. Wang, F. Wu, X. Chen, X. Wan, Y. Wu, X. Liu, Y. Li, F. Zhu, L. He, J. Chen, Y. Zhang and J. Jiao, Microbiome, 2021, 9, 185 CrossRef CAS PubMed.
- J. Duan, Y. Song, X. Zhang and C. Wang, Front. Physiol., 2021, 12, 646491 CrossRef PubMed.
- H.-J. Zhang, X. Gao, X.-F. Guo, K.-L. Li, S. Li, A. J. Sinclair and D. Li, Clin. Nutr., 2021, 40, 4538–4550 CrossRef CAS PubMed.
- B. Wang, L. Wu, J. Chen, L. Dong, C. Chen, Z. Wen, J. Hu, I. Fleming and D. W. Wang, Signal Transduction Targeted Ther., 2021, 6, 94 CrossRef CAS PubMed.
- S. T. Tan, T. Ramesh, X. R. Toh and L. N. Nguyen, Prog. Lipid Res., 2020, 80, 101068 CrossRef CAS PubMed.
- L. S. Huang, N. D. Hung, D. E. Sok and M. R. Kim, Lipids, 2010, 45, 225–236 CrossRef CAS PubMed.
- N. D. Hung, D. E. Sok and M. R. Kim, et al. , Inflammation research: official journal of the European Histamine Research Society, 2012, 61, 473–483 CAS.
- G. Chocian, A. Chabowski, M. Zendzian-Piotrowska, E. Harasim, B. Łukaszuk and J. Górski, Mol. Cell. Biochem., 2010, 340, 125–131 CrossRef CAS PubMed.
- N. Turner, G. M. Kowalski, S. J. Leslie, S. Risis, C. Yang, R. S. Lee-Young, J. R. Babb, P. J. Meikle, G. I. Lancaster, D. C. Henstridge, P. J. White, E. W. Kraegen, A. Marette, G. J. Cooney, M. A. Febbraio and C. R. Bruce, Diabetologia, 2013, 56, 1638–1648 CrossRef CAS PubMed.
- K. Eisinger, G. Liebisch, G. Schmitz, C. Aslanidis, S. Krautbauer and C. Buechler, Int. J. Mol. Sci., 2014, 15, 2991–3002 CrossRef PubMed.
- M. Park, D. Wu, T. Park, C. S. Choi, R. K. Li, K. K. Cheng, A. Xu and G. Sweeney, Am. J. Physiol.: Endocrinol. Metab., 2013, 305, E795–E804 CrossRef CAS PubMed.
- S.-S. Im, H. Y. Park, J. C. Shon, I.-S. Chung, H. C. Cho, K.-H. Liu and D.-K. Song, PLoS One, 2019, 14, e0213285 CrossRef CAS PubMed.
- J. C. Nelson, X.-C. Jiang, I. Tabas, A. Tall and S. Shea, Am. J. Epidemiol., 2006, 163, 903–912 CrossRef PubMed.
- X. C. Jiang, F. Paultre, T. A. Pearson, R. G. Reed, C. K. Francis, M. Lin, L. Berglund and A. R. Tall, Arterioscler., Thromb., Vasc. Biol., 2000, 20, 2614–2618 CrossRef CAS PubMed.
- M. Darabi and A. Kontush, Biochimie, 2016, 120, 81–86 CrossRef CAS PubMed.
- M. R. Flock and P. M. Kris-Etherton, Curr. Opin. Clin. Nutr. Metab. Care, 2013, 16, 133–140 CrossRef CAS PubMed.
- D. J. Baer and J. A. Novotny, Am. J. Clin. Nutr., 2019, 109, 269–275 CrossRef PubMed.
- R. P. Mensink and O. World Health, Effects of saturated fatty acids on serum lipids and lipoproteins: a systematic review and regression analysis, World Health Organization, Geneva, 2016 Search PubMed.
- J. Praagman, E. A. de Jonge, J. C. Kiefte-de Jong, J. W. Beulens, I. Sluijs, J. D. Schoufour, A. Hofman, Y. T. van der Schouw and O. H. Franco, Arterioscler., Thromb., Vasc. Biol., 2016, 36, 2011–2018 CAS.
- J. Praagman, J. W. Beulens, M. Alssema, P. L. Zock, A. J. Wanders, I. Sluijs and Y. T. van der Schouw, Am. J. Clin. Nutr., 2016, 103, 356–365 Search PubMed.
- S. Hlais, D. El-Bistami, B. El Rahi, M. A. Mattar and O. A. Obeid, Lipids, 2013, 48, 853–861 CrossRef CAS PubMed.
- P. Pérez-Martínez, A. García-Ríos, J. Delgado-Lista, F. Pérez-Jiménez and J. López-Miranda, Curr. Pharm. Des., 2011, 17, 769–777 CrossRef PubMed.
- P. Brites, H. R. Waterham and R. J. Wanders, Biochim. Biophys. Acta, 2004, 1636, 219–231 CrossRef CAS PubMed.
- E. Nguma, S. Yamashita, K. Kumagai, Y. Otoki, A. Yamamoto, T. Eitsuka, K. Nakagawa, T. Miyazawa and M. Kinoshita, ACS Omega, 2021, 6, 3140–3148 CrossRef CAS PubMed.
- S. Yamashita, M. Hashimoto, A. M. Haque, K. Nakagawa, M. Kinoshita, O. Shido and T. Miyazawa, Lipids, 2017, 52, 575–585 CrossRef CAS PubMed.
- S. Mawatari, S. Ohara, Y. Taniwaki, Y. Tsuboi, T. Maruyama and T. Fujino, Parkinsons Dis, 2020, 2020, 2671070 Search PubMed.
- L. Ding, L. Zhang, H. Shi, C. Xue, T. Yanagita, T. Zhang and Y. Wang, J. Funct. Foods, 2020, 66, 103824 CrossRef CAS.
- M. Suzuki, M. Yoshioka, Y. Ohno and Y. Akune, Sci. Rep., 2018, 8, 12030 CrossRef PubMed.
- S. Venn-Watson, M. Baird, B. Novick, C. Parry and E. D. Jensen, PLoS One, 2020, 15, e0230769 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available: Tables S1 and S2. See DOI: https://doi.org/10.1039/d2mo00202g |
| ‡ Marine Microbial Ecology Laboratory, Biology Program, New York University Abu Dhabi, Abu Dhabi 129188, United Arab Emirates. |
| This journal is © The Royal Society of Chemistry 2023 |