
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceEffect of shape, gluten, and mastication effort on in vitro starch digestion and the predicted glycemic index of pasta
Xinying
Suo
ab,
Anna
Baggio
 c,
Nicoletta
Pellegrini
c,
Silvia
Vincenzetti
a and
Elena
Vittadini
*a
c,
Nicoletta
Pellegrini
c,
Silvia
Vincenzetti
a and
Elena
Vittadini
*a
aSchool of Biosciences and Veterinary Medicine, University of Camerino, via Gentile III da Varano, 62032, Camerino, MC, Italy. E-mail: elena.vittadini@unicam.it; Tel: +39 0737 403209
bSchool of Food and Bioengineering, Collaborative Innovation Centre of Food Production and Safety, Zhengzhou University of Light Industry, Zhengzhou, China
cDepartment of Agricultural, Food, Environmental and Animal Sciences, University of Udine, via Sondrio 2/A, Udine, 33100, Italy
First published on 15th December 2023
Abstract
Gluten-containing (GC) and gluten-free (GF) pasta consumption has been growing in recent years. The market offers a wide variety of pasta types, with differences in shape and formulation that influence the mastication process and, consequently, their nutritional behaviors (i.e. starch digestibility and glycemic response). This study investigated the effect of shape, gluten, and structural breakdown on in vitro starch digestibility and predicted the glycemic index (pGI) of GC and GF penne, spaghetti, and risoni. Pasta was cooked and minced to mimic short, intermediate, and long mastication efforts. Short mastication led to a higher number of big particles than intermediate and long mastications for all pasta samples, which was reflected in the different starch digestibility and pGI patterns. Multivariate analysis of variance showed that the three studied factors differently affected the in vitro starch digestion of pasta. Mastication effort, shape, and their interaction mainly affected the starch digestion rate and pGI. Gluten was the major factor in affecting the amount of digested starch. The results suggested that small shapes (i.e. risoni), the presence of gluten, and short mastication effort led to a lower pGI. The findings will be useful for the development of pasta products tailored to fulfill the needs of specific consumers following a rational food design approach.
1. Introduction
The global pasta market size has been steadily growing in recent years (from USD 43.63 billion in 2021 to USD 46.84 billion in 2022) and is forecast to reach USD 77.83 billion by 2029, exhibiting a compound annual growth rate of 7.52% (2022–2029). The global COVID-19 pandemic significantly impacted the global pasta market, which experienced a 7.84% growth in 2020 as compared to 2019.1 The great success of pasta is related to its affordable price, easiness to cook, and long shelf-life, and the rising adoption of Western food habits is expanding pasta consumption all over the world. The pasta market is dominated by durum wheat-based products but, in recent years, the gluten-free pasta segment, developed to respond to meet the needs of celiac individuals, has significantly expanded to represent about 2% of the global pasta market (US$ 1.1 billion in 2022 with a compound annual growth rate of 4.5% [2022–2032]).2 Pasta is a staple food for a large part of the world's population. Pro capita pasta consumption varies in different parts of the world, reaching a maximum of 23 kg per year in Italy and around 8–11 kg in other European and American countries.3 It plays a key role in the daily diet and, therefore, it is important to understand the factors involved with the release of its nutrients, mainly starch, during digestion.During digestion, food matrix breakdown plays a pivotal role in how nutrients and bioactive compounds are made available for absorption in the human body, therefore regulating their concentration in the blood and their utilization in peripheral tissues.4,5 Food matrix breakdown depends not only on the mastication effort (time of mastication) but is also related to the product's macrostructural and microstructural attributes, as indicated in previous studies.6–8 Pasta macrostructural attributes are easily modulated by the use of different dies at the end of the extrusion process, which leads to the formation of pasta pieces with a large variety of shapes. At a microscopic scale, the presence or lack of a gluten network can importantly modify structural attributes in gluten-containing (semolina-based, GC) or gluten-free (e.g. maize- and rice-based, GF) products.
It was previously reported that pasta macrostructural attributes (the shape of the pasta) influenced both the mastication effort and the shape and size of the pieces present in boluses ready to be swallowed.6,9,10 In particular, penne-shaped pasta is masticated for a longer time than spaghetti,9,10 and further significantly longer than risoni in both GC and GF products.9 The resulting boluses were also composed of residual pieces with markedly different particle size distributions. Risoni were characterized by more pieces of small particle size, while spaghetti and penne led to the generation of more pieces of large particle size in an actual oral processing experiment.9,10 Furthermore, the size reduction of masticated pieces was found to increase the blood glucose release of cooked spaghetti compared to their intact strands.11 A relatively lower glycemic response was found in consuming long pasta (spaghetti) compared with short pasta (penne) in an in vivo study, indicating a possible influence of different extrusion processes on the structural organization of the pasta matrix, which could influence starch digestibility.10
The presence/absence of a gluten network is often taken as the most important pasta microstructural attribute and it has been recognized to affect both the pasta quality and the nutrient bioavailability.12–14 In a study of pasta oral processing, it was found that in pasta with the same macroscopic shape, gluten increased the chewing effort (chew number), but only marginally the particle size distribution of pieces present in ready-to-swallow boluses.9
It is here hypothesized that the macro- and micro-structural attributes of pasta, as well as the mastication effort, influence its structural breakdown, resulting in boluses composed of different sized pieces that, in turn, may play important roles in digestion and nutrient uptake. To the best of the authors’ knowledge, the effect of pasta shape, gluten, and in vitro structural breakdown on starch digestion and predicted glycemic response has not yet been fully studied. This study aims, therefore, to investigate the effect of macro- (shape) and micro- (presence/absence of a gluten network) structure, as well as the degree of structural breakdown (mastication effort), on in vitro starch digestion and the predicted glycemic index of pasta.
2. Materials and methods
2.1 Pasta samples
Commercial GF (90% corn flour, 10% rice flour, water; Massimo Zero, Merano, Italy) and GC (durum wheat semolina, water; Barilla, Parma, Italy) penne, spaghetti, and risoni were used in this study. All products were cooked in boiling deionized water (pasta![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :water = 1:10), without salt, for the cooking time suggested by producers as previously reported.9 After cooking, pasta was quickly drained and cooled at room temperature for 3 min. The moisture content of the cooked pasta was measured (after drying at 105 °C to constant weight) to standardize the starch digestibility based on the dry matter. The water content was the following: 51.5% for GF penne, 48.5% for GC penne; 60.7% for GF spaghetti, 56.7% for GC spaghetti; 63.8% GF risoni, for 70% GC risoni.
:water = 1:10), without salt, for the cooking time suggested by producers as previously reported.9 After cooking, pasta was quickly drained and cooled at room temperature for 3 min. The moisture content of the cooked pasta was measured (after drying at 105 °C to constant weight) to standardize the starch digestibility based on the dry matter. The water content was the following: 51.5% for GF penne, 48.5% for GC penne; 60.7% for GF spaghetti, 56.7% for GC spaghetti; 63.8% GF risoni, for 70% GC risoni.
2.2 Structural breakdown of cooked pasta
Cooked pasta samples were subjected to three in vitro structural breakdowns to represent short, intermediate, and long mastication efforts. The intermediate mastication effort aimed to reproduce the particle size distribution found in ready-to-swallow boluses produced by consumers measured in an oral processing experiment.9Pasta structural breakdown was carried out in vitro in order to control and standardize the amylase content in ready-to-swallow boluses. Preliminary tests were carried out first to identify the chopping conditions that best replicated the particle size distribution of pieces present in ready-to-swallow boluses and set as intermediate mastication.9 Cooked penne and spaghetti (100 g) were minced in a food processor bowl (1.2 L capacity; Magimix Cook Expert, 18900IT; MK2shop, Udine, Italy) for different times (5, 14, 15, 17, 19, 20, 22, 25, and 40 s for penne; 3, 5, 6, 8, 10, 12, and 16 s for spaghetti). Cooked risoni (5 g) were placed into a 10 mL beaker (2.5 cm diameter) and manually chopped (using a 2 cm blade knife) several times (0, 5, 6, 7, and 8 cuttings). Minced cooked pasta pieces were then characterized as previously described.9 Briefly, 5 g of bolus was randomly taken and the constituent pieces were separated and placed on a flatbed scanner (Canon CanoScan Lide 400, USA) to obtain a TIFF image (RGB mode, 600 ppi resolution). Collected images were analyzed using Image J software (IJ 1.46r, NIH) by setting the scale and smoothing, and converting to 8-bit images, in order to count the number of fragments and to measure their particle sizes (area, mm2). Three replicates of each pasta type were analyzed for each mincing time considered. Data were classified into 9-dimensional classes (1: ≤9.9; 2: 10.0–19.9; 3: 20.0–29.9; 4: 30.0–39.9; 5: 40.0–49.9; 6: 50.0–59.9; 7: 60.0–69.9; 8: 70.0–79.9; and 9: ≥80 mm2). The particle size distributions for each pasta type were compared with those of oral processing to identify the most similar distribution to masticated bolus.9 Intermediate mastication distributions were identified and set to correspond to 19 s chopping in a food processor for penne (GF and GC), 8 s and 6 s for GF and GC spaghetti, respectively, and 6 manual choppings for risoni (GF and GC).
Short mastication was arbitrarily set to be equal to 5 s and 3 s chopping in the food processor for, respectively, penne (GF and GC) and spaghetti (GF and GC), while risoni was not subjected to cutting. Particle size distributions of short mastication pasta samples were also measured.
Long mastication was set to correspond to the complete mashing of the pasta structure and this was achieved by passing 50 g of cooked product through a meat grinder (ARTUS T25, Reber Srl, Italy), as is commonly done in the determination of in vitro starch digestion.15–18
2.3 In vitro starch digestion and predicted glycaemic index
Pasta samples with particle size distributions corresponding to short, intermediate, and long mastication were subjected to the in vitro starch digestion procedure, as previously reported with slight modifications, to determine the amount of released glucose.19 Pepsin–guar gum solution (pH = 3) was prepared to simulate gastric phase as described in the method including pepsin (5 mg mL−1; P-7000, 444 U mg−1, Sigma) and guar gum (5 mg mL−1; G-4129, Sigma). The enzyme mixture was prepared from pancreatin (P-7545, 8× USP, Sigma, trypsin activity 6 U mg−1), amyloglucosidase (A7095, Sigma), and invertase (I4504, Sigma). Pancreatin was suspended in water at 150 mg mL−1 and, after 10 min of magnetic stirring, was centrifuged at 1500g at 4 °C for 5 min. Afterward, three-fourths of the cloudy supernatant were taken out to further mix with amyloglucosidase (0.04 mL mL−1 in the final mixture) and invertase (0.562 mg mL−1 in the final mixture) to achieve the final enzyme mixed suspension. All enzyme solutions were freshly prepared.Five grams of pasta of each mastication effort were statically incubated for 30 min with 10 mL of pepsin–guar gum solution in a water bath at 37 °C. Later, 10 mL of 0.25 M sodium acetate and 5 glass marbles (∅ 15 mm) were added, and then 5 mL of the enzyme mixture was added for further incubation with shaking (37 °C, 160 strokes per min, 120 min). Every 15 min from 0 to 120 min, 0.1 mL of each sample was collected, immediately mixed with 0.4 mL of absolute ethanol to stop the hydrolysis reaction, and diluted with water (1:10). The diluted samples were vortexed and centrifuged (3000g, 10 min) to obtain the supernatant for glucose measurement, which was determined using a D-glucose assay kit (K-GLUC, Megazyme, Bray, Ireland). A blank, containing water rather than all the enzymes used for digestion, was also carried out. The starch digested at each time interval was calculated based on the amount of glucose multiplied by the conversion factor of 0.9 and was expressed in g per 100 g dry cooked pasta for the evaluation of starch digestibility and g per 100 g cooked pasta to calculate the predicted glycemic index (pGI). The pGI was calculated using white bread digestion as a reference. The in vitro digestion of each sample was conducted in three replicates.
The digested starch values of each mastication level of each pasta sample (g per 100 g dry cooked pasta) were fitted to the Box–Lucas model (eqn (1)),20 using OriginPro 9.1 (OriginLab Corporation, Northampton, MA, USA):
| Ct = C∞ (1 − e−kt) | (1) |
Digested starch values of pasta (g per 100 g cooked pasta) and white bread (g per 100 g bread) were used for calculating the hydrolysis index (HI). The HI was derived from the ratio between the area under the hydrolysis curve (0–120 min) and the corresponding area of white bread expressed as a percentage over the same digestion time. The pGI for each mastication level of each pasta sample was calculated based on the following formula: pGI = 8.198 + 0.862 HI.22
2.4 Statistical analyses
Data were analyzed using IBM SPSS Statistics 22 (IBM Corporation, New York, USA). All the results were normally distributed as verified with the Shapiro–Wilk test (p < 0.05) and expressed as mean ± standard deviation (SD) of three replicates. Significant differences of starch digested at 90 min (C90), 120 min (C120), infinite time (C∞) and the digestibility rate constant (k), and the predicted glycemic index (pGI) between samples were identified through one-way analysis of variance (ANOVA, p < 0.05) followed by Duncan's test. A t-test (p < 0.05) was used to verify the significant difference of C90, C120, C∞, k, and pGI between GF and GC pasta with the same shape. Variance homogeneity was previously verified according to the Levene test (p < 0.05). To better understand the correlation between the pGI and C values, Pearson's correlation coefficient was run. A multivariate analysis of variance (MANOVA) based on Pillai's Trace test was run with shape, gluten, and mastication level as fixed factors (at a significance level of α = 0.05) to assess the influence of pasta shape, gluten, and mastication level on C90, k, and pGI.3. Results
3.1 Particle size distribution of the chopped pasta
Particle size distribution analysis was possible for short and intermediate mastication of pasta but not for long mastication as these samples were completely mashed at the end of the long oral processing simulation. In general, short mastication led to a higher number (p < 0.05) of big particles than the intermediate mastication for all pasta shapes (Fig. 1). Sixty-nine percent of GF penne, 90% of GC penne, 48% of GF spaghetti, and 40% of GC spaghetti pieces had an area larger than 80 mm2 when short mastication was simulated. In contrast, short mastication of GF and GC risoni generated more than three-quarters of pieces in the ranges 20.0–29.9 mm2 and 30.0–39.9 mm2, respectively. Intermediate mastication of all samples generated a prevalence of particle size pieces between the first and fourth classes, which means a range between 0 and 39.9 mm2. For GF and GC penne, a reduction in both length and width was observed, producing heterogeneously shaped fragments, while GF and GC spaghetti were shortened in length resulting in short spaghetti strands.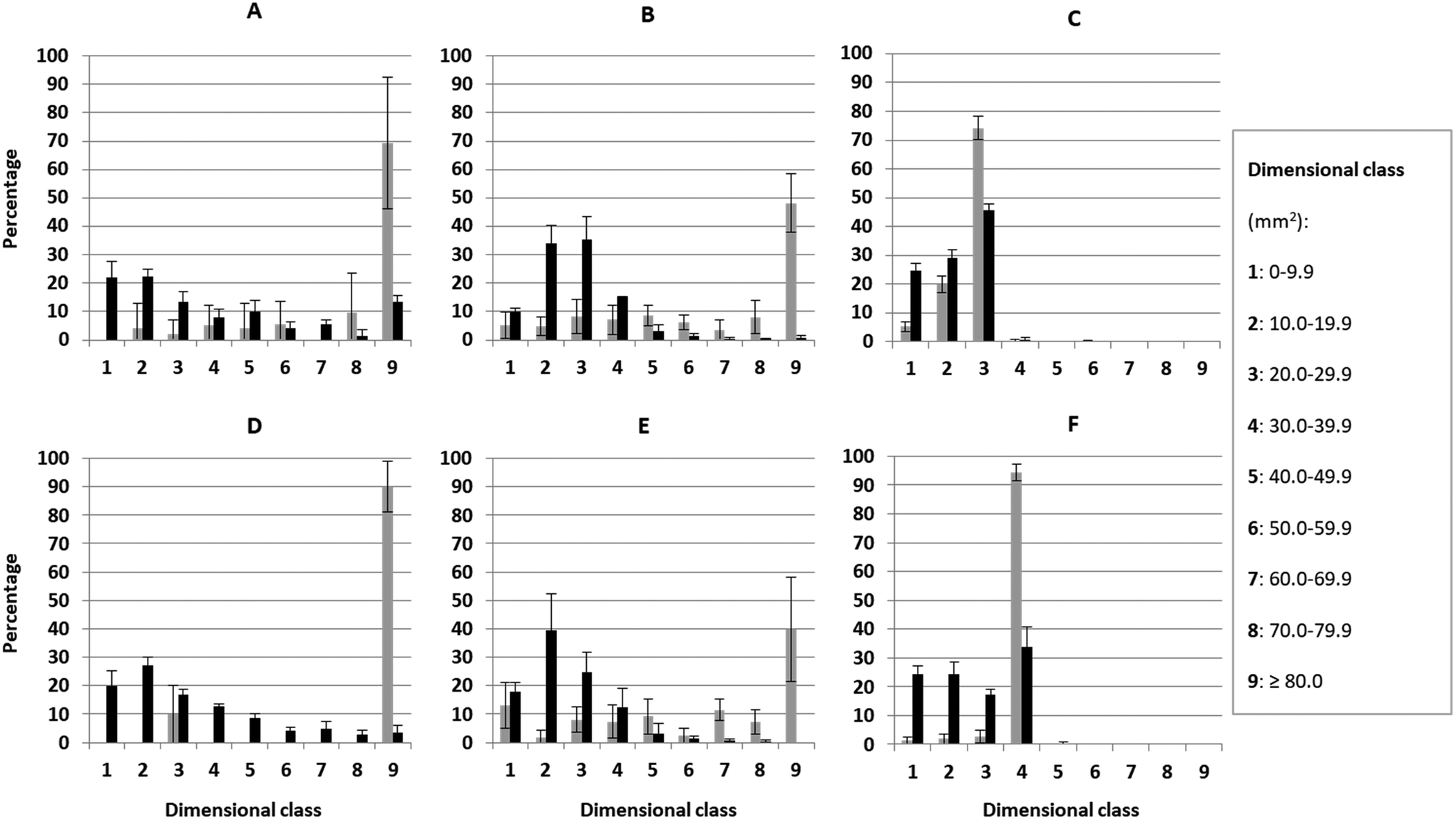 | ||
| Fig. 1 The particle size distribution (mean ± SD, n = 3) of GF (A: penne; B: spaghetti; and C: risoni) and GC (D: penne; E: spaghetti; and F: risoni) cooked pasta with short (gray bars) and intermediate (black bars) mastication. | ||
3.2 In vitro starch digestion of minced pasta
In vitro starch digestion of different GF and GC pasta shapes was examined by fitting the digestion curves to the Box–Lucas model (R2 ≥ 0.97 for all samples, Table 1 and Fig. 2). The rate of starch digestion (k) revealed a different pattern as a function of the mastication effort (F = 61.8, p < 0.001; Table 2), the shape of pasta (F = 81.9, p < 0.001), and their interaction (F = 7.6, p < 0.001). k increased with the extent of mastication with a steep increase in the starch digestion curves in the first 15 min for all pasta samples (Fig. 2). Moreover, at the same mastication effort, k differed among the three pasta shapes (Table 1). Following the digestion, the amount of digested starch increased progressively until 120 min following a first-order kinetic except for the curve of short masticated GC penne, which had a very low k (0.012 ± 0.001) and an unreliable calculated C∞ (87.9 ± 9.0 g per 100 g dry cooked pasta). In general, the trend showed that starch digested at C90 was lower than that at C120, which, in turn, was similar to that at C∞. This was not true for GC penne at short mastication effort meaning that the starch digestion cannot be considered near completion at 120 min. Except for risoni at short and intermediate mastication, the amount of starch digested at C90, C120, and C∞ was higher in GF pasta than that in their GC counterparts (Table 1), suggesting the main role of gluten on the starch digestibility as a single factor (F = 76.0, p < 0.001; Table 2) and as interaction with pasta shape (F = 63.4, p < 0.001) and mastication (F = 9.2, p < 0.001). The amount of digested starch was also influenced by mastication (F = 27.0, p < 0.001), which positively contributed to starch digestion when the time of mincing was increased.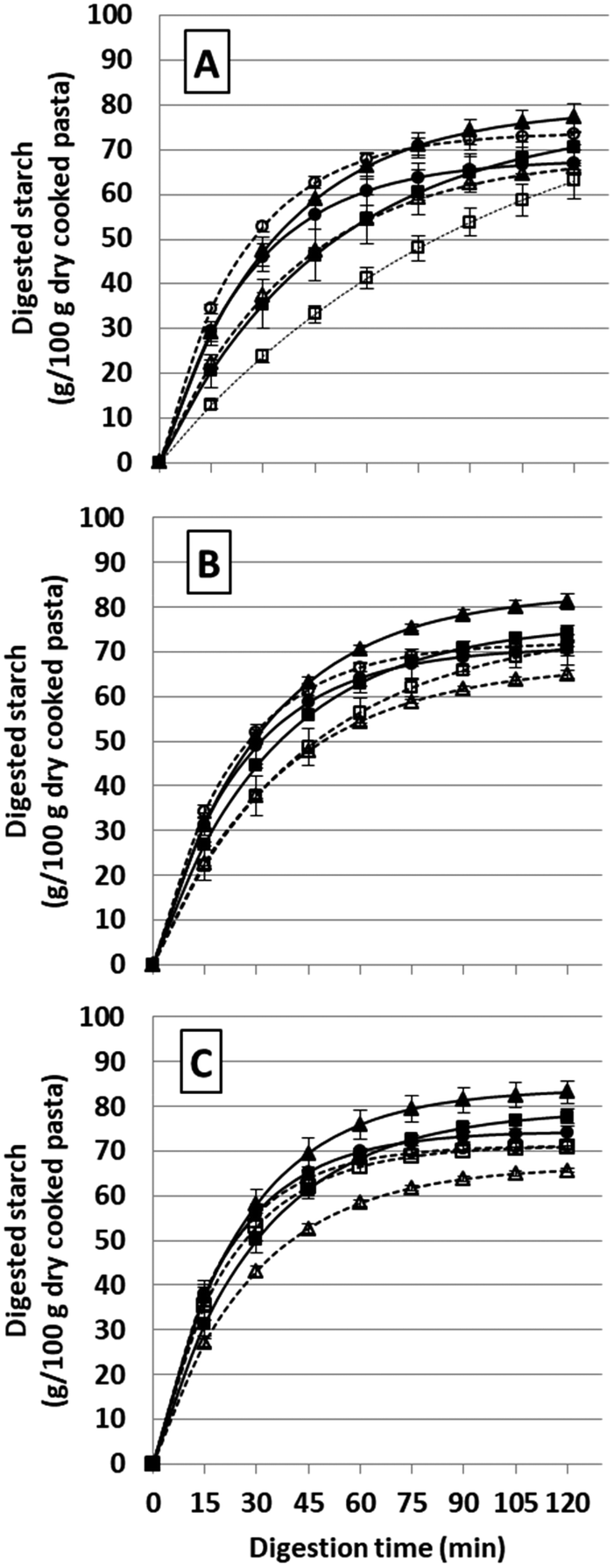 | ||
| Fig. 2 Pasta starch (g per100 g dry cooked pasta) digested at different mastication efforts (A: short; B: intermediate; and C: long). Black squares: GF penne; white squares: GC penne; black triangles: GF spaghetti; white triangles: GC spaghetti; black circles: GF risoni; and white circles: GC risoni. Lines represent the fitting of the Box–Lucas model. Data are expressed as mean ± SD (n = 3). | ||
| Shape | Mastication effort | C 90 | C 120 | C ∞ | k | |
|---|---|---|---|---|---|---|
| Data are expressed as mean ± SD. For each formulation and mastication effort, different letters mean significant differences of the same digestibility parameter between different pasta shapes (GF: lowercase; GC: uppercase) (p < 0.05, Duncan's test, n = 3). For each pasta shape and mastication level, asterisks mean significant differences between the same digestibility parameter of the GF and GC counterparts (*: p < 0.05; **: p < 0.01; ***: p < 0.001, t-test, n = 3). For each pasta and digestibility parameter, different superscript letters mean significant differences between mastication efforts (GF: lowercase; GC: uppercase) (p < 0.05, Duncan's test, n = 3). | ||||||
| GF | Penne | Short | 66.6 ± 4.7 a b | 70.7 ± 3.3 b b | 78.1 ± 2.6 a a | 0.020 ± 0.005 b*b |
| Intermediate | 71.5 ± 1.9 b ab | 74.7 ± 1.7 b ab | 76.5 ± 2.3 b a | 0.029 ± 0.002 b a | ||
| Long | 75.3 ± 1.9 b*a | 78.8 ± 2.6 ab**a | 79.4 ± 2.3 b* a | 0.033 ± 0.005 b*a | ||
| Spaghetti | Short | 73.2 ± 2.0 a***c | 79.6 ± 3.3 a*a | 79.5 ± 3.6 a**a | 0.030 ± 0.004 a b | |
| Intermediate | 78.6 ± 1.1 a***b | 83.4 ± 3.6 a**a | 83.2 ± 2.4 a**a | 0.032 ± 0.003 b b | ||
| Long | 83.3 ± 2.7 a***a | 85.2 ± 2.3 a***a | 83.9 ± 2.4 a***a | 0.039 ± 0.002 ab a | ||
| Risoni | Short | 64.3 ± 5.6 a b | 70.3 ± 1.3 b a | 67.9 ± 2.5 b*b | 0.038 ± 0.002 a b | |
| Intermediate | 67.2 ± 2.2 c ab | 72.8 ± 4.4 b a | 71.2 ± 3.5 b ab | 0.039 ± 0.004 a b | ||
| Long | 73.8 ± 0.1 b***a | 75.6 ± 1.2 b a | 74.4 ± 1.6 c*a | 0.047 ± 0.005 a a | ||
| GC | Penne | Short | 62.4 ± 4.6 B B | 68.0 ± 4.8 A A | 87.9 ± 9.0 A A | 0.012 ± 0.001 C C |
| Intermediate | 66.9 ± 4.0 A AB | 72.0 ± 0.5 A A | 76.7 ± 6.7 A AB | 0.023 ± 0.006 B B | ||
| Long | 70.0 ± 1.7A A | 71.2 ± 1.1 A A | 71.3 ± 0.5 A B | 0.046 ± 0.006 A A | ||
| Spaghetti | Short | 61.7 ± 0.7 B B | 67.6 ± 0.9 A A | 69.1 ± 1.3 B A | 0.026 ± 0.002 B B | |
| Intermediate | 61.4 ± 0.1 B B | 67.9 ± 1.4 B A | 67.4 ± 1.4 B AB | 0.028 ± 0.002 B B | ||
| Long | 63.6 ± 0.7 B A | 66.3 ± 0.5 A A | 66.7 ± 0.5 B B | 0.035 ± 0.001 B A | ||
| Risoni | Short | 72.0 ± 3.6 A A | 72.7 ± 1.4 A A | 74.0 ± 2.6 B A | 0.042 ± 0.003 A A | |
| Intermediate | 70.4 ± 2.3 A A | 72.8 ± 1.8 A A | 72.1 ± 0.3 AB A | 0.043 ± 0.002 A A | ||
| Long | 69.8 ± 0.7 A A | 71.0 ± 4.0 A A | 71.2 ± 1.0 A A | 0.051 ± 0.007 A A | ||
| C 90 | k | pGI | ||
|---|---|---|---|---|
| *: p < 0.05; **: p < 0.01; ***: p < 0.001; and ns: not significant. | ||||
| Shape | F | 1.8 | 81.9 | 337.9 |
| Sig. | ns | *** | *** | |
| Gluten | F | 76.0 | 0.2 | 93.2 |
| Sig. | *** | ns | *** | |
| Mastication | F | 27.0 | 61.8 | 85.5 |
| Sig. | *** | *** | *** | |
| Shape × Gluten | F | 63.4 | 5.5 | 13.2 |
| Sig. | *** | ** | *** | |
| Gluten × Mastication | F | 9.2 | 4.6 | 1.2 |
| Sig. | *** | * | ns | |
| Shape × Mastication | F | 2.0 | 7.6 | 18.6 |
| Sig. | ns | *** | *** | |
| Shape × Gluten × Mastication | F | 2.5 | 4.6 | 4.9 |
| Sig. | ns | ** | ** | |
3.3 Predicted glycemic index
The pGI progressively increased by increasing mastication effort (F = 85.5, p < 0.001; Table 2), especially for penne, increasing from below 55 with short mastication to nearly 70 with long mastication (Fig. 3). GF spaghetti showed low pGI (<55) at short and intermediate mastication and medium pGI (55 < GI < 69) at long mastication time, while the pGI of GC spaghetti and risoni remained low at all mastication efforts. Pasta shape was found to be the main factor influencing the pGI (F = 337.9, p < 0.001; Table 2) as, independently from the mastication time, the penne pGI was higher than the spaghetti pGI, which, in turn, was higher than the risoni pGI. Moreover, the pGI of GC pasta was generally lower than its GF counterparts, suggesting an important role of gluten in influencing the predicted glycaemic index (F = 93.2, p < 0.001).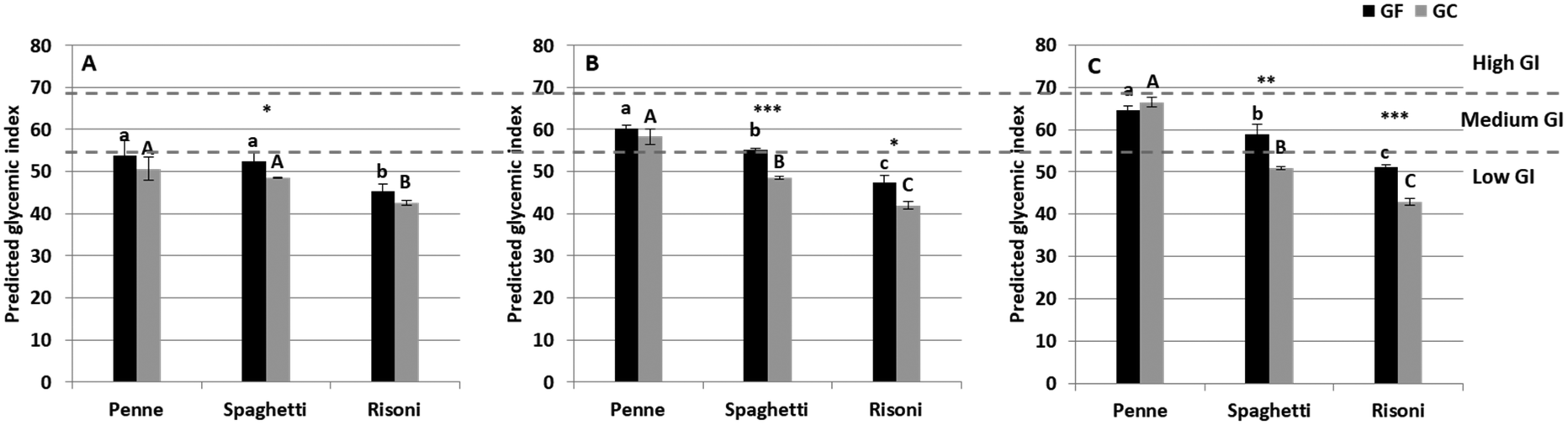 | ||
| Fig. 3 Predicted glycemic index (mean ± SD) of GF (black) and GC (gray) pasta for short (A), intermediate (B), and long (C) mastication times. Different letters in each subfigure mean significant differences between pasta with different shapes and the same formulation (lowercase for GF, uppercase for GC) (p < 0.05, Duncan's test, n = 3). Asterisk means significant differences between GF and GC pasta with the same shape at the same mastication level (* = p < 0.05, ** = p < 0.01, *** = p < 0.001, and t-test, n = 3). | ||
4. Discussion
Food oral processing is the first step of digestion. It is characterized by a large inter-individual variability that produces boluses with pieces of different particle sizes that may affect the digestion rate of nutrients (i.e. the starch digestion rate and glycemic response).5,6,9,23–25 As pasta is a staple food widely consumed in different shapes and formulations, the effects of its macroscopic (shape) and microscopic (gluten network) structure and the degree of structural breakdown related to different mastication efforts on starch digestion were studied using an in vitro digestion protocol and the predicted GI. The three studied factors affected differently the in vitro starch digestion of pasta. Mastication effort, shape, and their interaction mainly affected the starch digestion rate (i.e., k) and the pGI. Gluten was the major factor in affecting the amount of digested starch.The different composition and structural organization of GF and GC pasta were expected to affect water uptake by the pasta matrix, leading to a different moisture content of the cooked products. Therefore, to highlight the effect of structure (macroscopic and microscopic) and mastication effort on the starch digestion rate of pasta with different shapes, with or without gluten, and differently masticated, it was necessary to dry the cooked pasta samples to remove the effect of different water content gained during cooking between GF and GC pasta. In contrast, the pGI was calculated on the wet weight of cooked pasta, because GI in vivo is measured on food as eaten, and is correlated with C90. This result was in agreement with the findings of Edwards et al.,21 who studied the validity of the Box–Lucas model to fit the starch digestion of starch-rich foods concluding that the extrapolated C∞ might not be appropriate to describe the behavior of some samples.
Regarding the effect of microscopic structure (i.e., gluten), the amount of digested starch (C90) was higher in GF penne and spaghetti than that in their GC counterparts at each mastication level. These findings underlined a very strong impact of gluten microstructure on starch digestion, supporting the extensively reported hypothesis that the three-dimensional gluten network formed during kneading of semolina-based pasta entraps gelatinized starch, and thereby, acts as a physical barrier to limit the enzyme hydrolysis of starch, contributing to a lower starch digestibility.13,14 Moreover, the GF pasta analyzed in our study was formulated with corn and rice, which are cereals known to have a higher starch digestibility than wheat.22,26
Shape had a major effect on the pGI independently from the gluten presence. Penne had the highest pGI, followed by spaghetti and risoni. The higher pGI of penne compared to spaghetti agreed with the results of the post-prandial in vivo glucose response measured by Vanhatalo et al.10 Besides the macroscopic and microscopic structure, the C90 and pGI were affected by the mastication level as the values progressively increased by increasing the mincing time for the three types of pasta, as expected. From short to intermediate to long mastication, the number of small particles progressively increased at the expense of larger particles, leading to a higher contact surface for the enzymatic activity and, consequently, to a higher amount of digested starch that caused, in turn, the rise of the pGI. The particle size was already reported to affect the starch digestibility and pGI of bread having the same formulation but different matrixes, such as roll bread and loaf bread.25 In particular, more extensively simulated bread mastication led to the formation of smaller particle sizes than a less fractured bolus.25 This result agrees with our finding, emphasizing that food structure and the oral mastication phase significantly affect the bolus particle size distribution. Previous studies on rice also revealed a positive correlation between the mastication degree, starch digestibility, and pGI, demonstrating that the dimension of rice particles affected the in vitro digestibility of starch with the smaller particles digested at a higher rate.24 Moreover, they proved by an in vivo study that the glycemic index of rice chewed 15 times was significantly lower than when masticated 30 times (GI 68 and 88, respectively).27 In contrast, risoni contradicted these results having the highest number of small particles but the lowest pGI. However, as previously reported, risoni were swallowed as almost not masticated pieces.9 This behavior might be related to the harder structure of risoni compared to penne and spaghetti, which might have limited the enzyme penetration and, consequently, the amount of digested starch. Moreover, risoni gained more water during cooking than penne and spaghetti, resulting in lower total starch content and, consequently, a lower pGI.
5. Conclusion
In this study, the starch digestibility and pGI of pasta having different shapes and formulations was investigated through three levels of mastication. Pasta shape was found to be the predominant factor in affecting the starch digestion rate and pGI. Moreover, the presence of an organized microstructure (i.e. the continuous gluten network with embedded gelatinized starch) reduced the starch digestion and the predicted glycemic response. A more extensive mastication effort, with consequent more extensive physical structural breakdown, facilitated starch digestion and induced a higher expected glycemic index. The present results suggest that small pasta like risoni, gluten-containing products, or less mastication effort can be a strategy to have a relatively lower expected glycemic index. The findings of this investigation will be useful for the development of pasta products tailored to fulfill the needs of specific consumers following a rational food design approach.Author contributions
Xinying Suo: conceptualization – equal, data curation – lead, formal analysis – lead, investigation – lead, visualization – equal, and writing – original draft – equal. Anna Baggio: writing – original draft – equal, data curation – equal, and formal analysis – equal. Nicoletta Pellegrini: conceptualization – equal, supervision – equal, and writing – review & editing – equal. Silvia Vincenzetti: supervision – equal, and writing – review & editing – equal. Elena Vittadini: conceptualization – equal, funding acquisition – lead, project administration – lead, supervision – equal, writing – review & editing – equal, and visualization – equal.Conflicts of interest
There are no conflicts to declare.References
- Fortune Business Insights TM, Pasta market size, share & COVID-19 impact analysis, by product type (dried, chilled, and canned), raw material (wheat and gluten-free), distribution channel (supermarkets/hypermarkets, convenience stores, online stores, and others), and regional forecast, 2022–2029. https://www.fortunebusinessinsights.com/pasta-market-102284 (2022). [Accessed on May 12th 2022].
- Future Market Insights. Gluten free pasta market. https://www.futuremarketinsights.com/reports/gluten-free-pasta-market ( 2022). [Accessed on May 12th 2022].
- Food Business News, Worldwide pasta consumption on the rise. ( 2018). [Accessed on June 1st 2022].
- E. Capuano and A. E. M. Janssen, Food matrix and macronutrient digestion, Annu. Rev. Food Sci. Technol., 2021, 12, 193–212 CrossRef CAS PubMed.
- J. Nadia, J. Bronlund, R. P. Singh, H. Singh and G. M. Bornhorst, Structural breakdown of starch-based foods during gastric digestion and its link to glycemic response: In vivo and in vitro considerations, Compr. Rev. Food Sci. Food Saf., 2021, 20, 2660–2698 CrossRef CAS PubMed.
- C. Hoebler, M. F. Devaux, A. Karinthi, C. Belleville and J. L. Barry, Particle size of solid food after human mastication and in vitro simulation of oral breakdown, Int. J. Food Sci. Nutr., 2000, 51, 353–366 CrossRef CAS PubMed.
- J. Chen, Food oral processing—A review, Food Hydrocolloids, 2009, 23, 1–25 CrossRef CAS.
- J. M. Aguilera, The food matrix: Implications in processing, nutrition and health, Crit. Rev. Food Sci. Nutr., 2019, 59, 3612–3629 CrossRef CAS PubMed.
- X. Suo, A. C. Mosca, N. Pellegrini and E. Vittadini, Effect of pasta shape and gluten on pasta cooking quality and structural breakdown during mastication, Food Funct., 2021, 12, 11577–11585 RSC.
- S. Vanhatalo, M. Dall'Asta, M. Cossu, L. Chiavaroli, V. Francinelli, G. Di Pede, R. Dodi, J. Närväinen, M. Antonini and M. Goldoni, Pasta structure affects mastication, bolus properties, and postprandial glucose and insulin metabolism in healthy adults, J. Nutr., 2022, 152, 994–1005 CrossRef PubMed.
- M. Petitot, C. Brossard, C. Barron, C. Larré, M.-H. Morel and V. Micard, Modification of pasta structure induced by high drying temperatures. Effects on the in vitro digestibility of protein and starch fractions and the potential allergenicity of protein hydrolysates, Food Chem., 2009, 116, 401–412 CrossRef CAS.
- A. Marti and M. A. Pagani, What can play the role of gluten in gluten free pasta?, Trends Food Sci. Technol., 2013, 31, 63–71 CrossRef CAS.
- J. A. Delcour, I. J. Joye, B. Pareyt, E. Wilderjans, K. Brijs and B. Lagrain, Wheat gluten functionality as a quality determinant in cereal-based food products, Annu. Rev. Food Sci. Technol., 2012, 3, 469–492 CrossRef CAS PubMed.
- W. Zou, M. Sissons, M. J. Gidley, R. G. Gilbert and F. J. Warren, Combined techniques for characterising pasta structure reveals how the gluten network slows enzymic digestion rate, Food Chem., 2015, 188, 559–568 CrossRef CAS PubMed.
- M. Petitot, J. Abecassis and V. Micard, Structuring of pasta components during processing: impact on starch and protein digestibility and allergenicity, Trends Food Sci. Technol., 2009, 20, 521–532 CrossRef CAS.
- G. Mamone, C. Nitride, G. Picariello, F. Addeo, P. Ferranti and A. Mackie, Tracking the fate of pasta (T. durum semolina) immunogenic proteins by in vitro simulated digestion, J. Agric. Food Chem., 2015, 63, 2660–2667 CrossRef CAS PubMed.
- R. Dodi, L. Bresciani, B. Biasini, M. Cossu, F. Scazzina, F. Taddei, M. G. D'Egidio, M. Dall'Asta and D. Martini, Traditional and non-conventional pasta-making processes: Effect on in vitro starch digestibility, Foods, 2021, 10, 921 CrossRef CAS PubMed.
- X. Suo, M. Dall'Asta, G. Giuberti, M. Minucciani, Z. Wang and E. Vittadini, The effect of chickpea flour and its addition levels on quality and in vitro starch digestibility of corn–rice-based gluten-free pasta, Int. J. Food Sci. Nutr., 2022, 1–10 CAS.
- K. N. Englyst, G. J. Hudson and H. N. Englyst, Starch analysis in food, Encycl. Anal. Chem., 2000, 4246–4262 Search PubMed.
- Y. Shen, D. Wu, V. Fogliano and N. Pellegrini, Rice varieties with a high endosperm lipid content have reduced starch digestibility and increased γ-oryzanol bioaccessibility, Food Funct., 2021, 12, 11547–11556 RSC.
- C. H. Edwards, N. Cochetel, L. Setterfield, N. Perez-Moral and F. J. Warren, A single-enzyme system for starch digestibility screening and its relevance to understanding and predicting the glycaemic index of food products, Food Funct., 2019, 10, 4751–4760 RSC.
- G. Giuberti, A. Gallo, C. Cerioli, P. Fortunati and F. Masoero, Cooking quality and starch digestibility of gluten free pasta using new bean flour, Food Chem., 2015, 175, 43–49 CrossRef CAS PubMed.
- M. L. Jalabert-Malbos, A. Mishellany-Dutour, A. Woda and M. A. Peyron, Particle size distribution in the food bolus after mastication of natural foods, Food Qual. Prefer., 2007, 18, 803–812 CrossRef.
- V. Ranawana, J. A. Monro, S. Mishra and C. J. K. Henry, Degree of particle size breakdown during mastication may be a possible cause of interindividual glycemic variability, Nutr. Res., 2010, 30, 246–254 CrossRef CAS PubMed.
- A. Aleixandre, Y. Benavent-Gil and C. M. Rosell, Effect of bread structure and in vitro oral processing methods in bolus disintegration and glycemic index, Nutrients, 2019, 11, 2105 CrossRef CAS PubMed.
- F. S. Atkinson, K. Foster-Powell and J. C. Brand-Miller, International tables of glycemic index and glycemic load values: 2008, Diabetes Care, 2008, 31, 2281–2283 CrossRef PubMed.
- V. Ranawana, M. K. S. Leow and C. J. K. Henry, Mastication effects on the glycaemic index: Impact on variability and practical implications, Eur. J. Clin. Nutr., 2014, 68, 137–139 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2024 |