
Probiotics enhance vitamin absorption in the intestine: influencing factors and potential mechanisms
Peiling
Hu
 ,
Han
Tan
,
Weiping
Xu
,
Shijian
Liu
,
Jiajia
Song
,
Xiaoyong
Chen
* and
Huayi
Suo
*
,
Han
Tan
,
Weiping
Xu
,
Shijian
Liu
,
Jiajia
Song
,
Xiaoyong
Chen
* and
Huayi
Suo
*
College of Food Science, Southwest University, Chongqing 400715, P. R. China. E-mail: birget@swu.edu.cn
First published on 24th June 2025
Abstract
Vitamins are essential for numerous bodily functions and metabolic regulation. The importance of probiotics in promoting intestinal absorption of vitamins has been confirmed, yet several factors influence the effectiveness of this promotion. Despite this, studies on the mechanisms by which probiotics facilitate this process remain limited. In this review, we discuss the potential mechanisms through which probiotics promote intestinal vitamin absorption. These include altering the composition of the intestinal microbiota, improving the intestinal environment, regulating intestinal pH, enhancing vitamin expression and activity, synthesizing transporter proteins, producing binding proteins, and generating postbiotics. Additionally, the promotion of intestinal vitamin absorption by probiotics may be influenced by digestive enzymes, vitamin transporter proteins, vitamin-binding proteins, and dietary lipids. Understanding the factors that influence how probiotics affect intestinal vitamin absorption and elucidating their underlying mechanisms of action are crucial for unlocking the full potential of probiotics and developing effective applications.
1. Introduction
Vitamins, which are primarily obtained through the diet, are essential for maintaining normal bodily functions. These substances can be classified into two categories based on their solubility: fat-soluble and water-soluble vitamins. Water-soluble vitamins include eight B vitamins and vitamin C, while fat-soluble vitamins include vitamins A, D, E, and K. Vitamin deficiency can lead to metabolic imbalances and decreased immunity, potentially resulting in malnutrition and an increased susceptibility to various functional diseases. For example, vitamin A deficiency can cause dry eye accompanied by night blindness, hyperkeratosis, and skin damage, and trigger infections.1 Vitamin B deficiency induces neurological deficiencies and immune and tumor disorders.2 Additionally, vitamin C deficiency can cause scurvy, which can be fatal in rare cases.3 Similarly, vitamin D deficiency can increase the risk of health issues including common cancers, multiple sclerosis, rheumatoid arthritis, hypertension, cardiovascular heart disease, and type I diabetes.4 Probiotics can synthesize vitamins in the gut and promote their absorption and utilization. Adequate vitamin intake, in turn, supports probiotic growth, creating a mutually beneficial relationship.Probiotics are live microorganisms that, when consumed in adequate quantities, provide health benefits to the hosts.5 Recent studies have demonstrated that probiotics not only produce vitamins required by the body but also promote their absorption in the intestinal tract. Probiotics achieve vitamin absorption through several mechanisms. These include regulating the intestinal microbial community,6 accelerating the release of vitamins from food, synthesizing transport proteins for vitamin transportation in the intestinal epithelium,7 and increasing the expression level of vitamin transporter proteins.8 However, it is important to note that the effects of probiotics are strain-specific, and different probiotics may not have the same effect on vitamin intestinal absorption. Currently, more research is needed to understand the effective strains, influencing factors, and mechanisms of action of probiotics for the intestinal absorption of vitamins.
This review describes the intestinal absorption pathways of vitamins, summarizes the enhancement of intestinal vitamin absorption through probiotics, and discusses the factors influencing this process. Finally, this review proposes potential mechanisms by which probiotics can promote intestinal vitamin absorption and explores future developments. These developments include the preparation of new probiotic supplements or foods to optimize vitamin intake and absorption, as well as investigations into how probiotics modulate the gut barrier function to improve vitamin permeability. A comprehensive understanding of the relationship between probiotics and intestinal vitamin absorption can facilitate the use of probiotics to prevent vitamin deficiencies.
2. Probiotics promote intestinal absorption of vitamins
2.1. Probiotics promote intestinal absorption of water-soluble vitamins
Water-soluble vitamins, including the B complex (B1, B2, B3, B5, B6, B7, B9, and B12) and vitamin C, are absorbed via active transport and passive diffusion in the gut (Fig. 1). Most water-soluble vitamins are typically ingested with food and enter the gastrointestinal (GI) tract in complex forms that are hydrolyzed by gastric acids, proteases, and digestive enzymes to produce binding proteins and free water-soluble vitamins. Human gut microbes actively synthesize B vitamins and provide them to neighboring bacteria through symbiotic relationships. A significant portion of the recommended vitamin B6 intake, along with approximately one-third of the recommended vitamin B9 and vitamin B12 intake, may be derived from the gut bacteria.9 At low concentrations of water-soluble vitamins in the gut, free vitamins bind to the carrier proteins. For example, the sodium-dependent transporter proteins sodium-dependent vitamin C transporter 1 (SVCT1) and sodium-dependent vitamin C transporter 2 (SVCT2) facilitate the absorption of vitamin C.10 Similarly, the thiamine transporter proteins thiamine transporter 1 (THTR-1) and thiamine transporter 2 (THTR-2) are utilized for the absorption of thiamine (vitamin B1).11 Other carrier proteins, including riboflavin transporter (RFVT), riboflavin transporter 2 (RFVT2), and riboflavin transporter 3 (RFVT3), are involved in the absorption of vitamin B2,12 while proton co-transporter proteins and anionic inverse transporter proteins play a role in the absorption of vitamin B3. These carrier proteins selectively transport water-soluble vitamins, thereby ensuring efficient absorption of each vitamin. Specifically, vitamin B12 binds to the glycoprotein intrinsic factor (IF) secreted by the gastric parietal cells to form a vitamin B12–IF complex that attaches to a specific receptor on the ileocecal terminal ileum and is then absorbed through receptor-mediated calcium-dependent active transport.13 | ||
| Fig. 1 Vitamins are absorbed in the small intestine. | ||
Water-soluble vitamins can also be absorbed via passive diffusion at higher concentrations. These vitamins do not require energy and are based on a concentration gradient. Most forms of vitamin B6 are absorbed in the jejunum and ileum via the passive diffusion pathway, and absorption is good even at high doses.14 However, only 1–2% of the oral dose of vitamin B12 is absorbed by passive diffusion.15 Water-soluble vitamins cannot be stored for extended periods in the body after absorption by the intestines and are tightly regulated by the kidneys.
Recent studies have highlighted the significant role of probiotics in promoting the absorption of water-soluble vitamins (Table 1). Each of the 12 participants consumed four portions of probiotic yogurt daily (125 g per portion, totaling 500 g), prepared with Streptococcus salivarius subsp. thermophilus, Lactobacillus acidophilus, and Lacticaseibacillus casei GG. Following yogurt consumption, the plasma vitamin B2 concentration significantly increased (p < 0.01).16 Female volunteers consumed 100 g day−1 of probiotic yogurt containing the starter cultures Streptococcus salivarius subsp. thermophilus (2 × 108 colony-forming units (CFU) per g) and Lactobacillus delbrueckii subsp. bulgaricus (1 × 107 CFU g−1), supplemented with the probiotic species Lacticaseibacillus casei (3.6 × 108 CFU g−1). The intervention lasted 2 weeks (T1–T2) at 100 g day−1, followed by 200 g day−1 for an additional 2 weeks (T2–T3). Plasma vitamin B1 levels increased significantly in both the probiotic group (p < 0.001) and the control group (p < 0.01) during the 200 g day−1 intake phase (T2–T3). Similarly, plasma flavin mononucleotide (FMN) levels showed significant elevations in both the probiotic group (p < 0.01) and the control group (p < 0.001) after the 100 g day−1 intake period (T1–T2). Additionally, free vitamin B2 concentrations also increased significantly in both groups (probiotic: p < 0.01; control: p < 0.001).17 Furthermore, Bifidobacterium adolescentis MB 239 elevated colonic vitamin B9 levels from 27 to 54 ng mL−1 in 48 hour fecal cultures, improving its absorption in the colon.18 Additionally, Limosilactobacillus reuteri CRL 1098 colonizes the intestine, produces vitamin B12, and influences the intestinal microbiota, thereby promoting the absorption of vitamin B12. Six-week-old pregnant BALB/c mice were administered Limosilactobacillus reuteri CRL 1098 (107 CFU per mouse per day) in aqueous suspension via oral gavage (0.5 mL daily). Pups from probiotic-treated dams exhibited significantly higher serum vitamin B12 levels (673.6 ± 34 pg mL−1) compared to those from B12-deficient dams (275.2 ± 38 pg mL−1).19 Therefore, probiotics can improve the absorption and overall status of water-soluble vitamins, contributing to better health outcomes.
| Probiotic species | Vitamin type | Animal model | Mechanisms involved | Ref. |
|---|---|---|---|---|
| Caco-2 cells: colon adenocarcinoma cells, SAMP6: senescence accelerated mouse prone 6, IBD: inflammatory bowel disease, VDR: vitamin D receptor, AMS: amylase, LPS: lipopolysaccharides, and TPS: terpene synthase. | ||||
| Streptococcus salivarius subsp. Thermophilus, Lacticaseibacillus casei | Vitamin B2 | 6 healthy males and six healthy females aged 25–36 years | Improve the B-vitamin contents | Ibrahim Elmadfa (2001)16 |
| Streptococcus salivarius subsp. Thermophilus, Lactobacillus delbrueckii subsp. Bulgaricus, Lacticaseibacillus casei | Vitamin B1, vitamin B2 | 33 healthy, nonsmoking, normocholesterolemic female participants aged between 22 and 29 years | Enhanced immune function and oxidant/antioxidant imbalance | Elisabeth Fabian (2008)17 |
| Bifidobacterium adolescentis | Vitamin B9 | Fecal culture experiments | Increase the folate concentration in the colonic environment and efficiently confer protection against inflammation and cancer | Anna Pompei (2007)18 |
| Limosilactobacillus reuteri | Vitamin B12 | Six-week-old pregnant BALB/c mice | Produced vitamin B12, alleviated intestinal inflammation, and regulated the structure of intestinal microbiota | V.C. Molina (2009)19 |
| Lacticaseibacillus rhamnosus | Vitamin D | Twelve 5 week-old specific-pathogen-free SAMP6 mice and in vitro Caco-2 cells | Increased expression of vitamin D transporter proteins NPC1L1 and CD36 promoted vitamin D intestinal absorption | Jing Cheng (2022)27 |
| Limosilactobacillus reuteri | Vitamin D | 127 adults: hypercholesterolemia (LDL-C > 3.4 mmol L−1), triglycerides <4.0 mmol L−1, BMI 22–32 kg m−2 | Increased intraluminal lactic acid production, increased synthesis of 7-DHC, and improved IBD symptomology, enhance VDR expression and activity | Mitchell L Jones (2013)28 |
| Lacticaseibacillus paracasei | Vitamin D | Male CD-1 mice aged 8 weeks | Probiotic-derived biosurfactants mediate vitamin D emulsification, which optimizes interfacial interactions with gut epithelial cells to potentiate absorptive efficacy | Ignazio Castagliuolo (2021)29 |
| Lactobacillus, Bifidobacterium | Multi-vitamins | Healthy and active Conger myriaster individuals of similar sizes, with an average body weight of 169.78 ± 3.25 g | Stimulated endogenous enzyme production in the host, significantly elevating the activities of AMS, LPS, and TPS, enhanced modulated intestinal microbiota homeostasis, improved microbial community richness and diversity, and increased the relative abundance of beneficial bacteria | Xinyu Zhao (2023)30 |
2.2. Probiotics promote intestinal absorption of fat-soluble vitamins
Probiotics have been shown to enhance the absorption of fat-soluble vitamins such as vitamins A, D, E, and K through various mechanisms. These vitamins are primarily absorbed via active transport and passive diffusion (Fig. 1). The parietal membrane proteins of enterocytes facilitate the absorption of fat-soluble vitamins at dietary doses, whereas at supplemental/drug doses, fat-soluble vitamins enter the enterocytes through passive diffusion.20 In the jejunum and ileum, the linear relationship between vitamin absorption and intestinal concentration suggests that intestinal absorption of fat-soluble vitamins is mediated by a passive, unpredictable diffusion mechanism that is neither energy-dependent nor carrier-mediated.21 Fat-soluble vitamins in food often bind to proteins to form complexes that are subjected to a combination of bile, pancreatic lipase, and enterolipase in the small intestine. Free fat-soluble vitamins can be released only after the complexes are broken down. Free vitamins need to be admixed into mixed micelles containing bile salts, pancreatic solubilization products, and other dietary lipids to be absorbed intestinally through the glycoprotein layer of the small intestinal villi.22 Parietal membrane transport proteins that are absorbed by the intestinal tract, such as scavenger receptor class B type I (SR-BI), cluster of differentiation 36 (CD36) and Niemann-Pick C1 Like 1 (NPC1L1), are usually available to mediate the absorption of vitamin A,23 vitamin D,24 vitamin E25 and vitamin K.26 Additionally, competition for SR-BI transport occurs between vitamins A and E and cholesterol.Probiotics have a positive effect on the absorption of fat-soluble vitamins. The presence of probiotic bacteria in the gut is essential for the effective absorption of these vitamins. Probiotics promote intestinal absorption of fat-soluble vitamins, as evidenced by several studies (Table 1). When undiluted Lacticaseibacillus rhamnosus GG culture supernatant (LCS) was administered to 5 week-old specific-pathogen-free senescence-accelerated mouse P6 (SAMP6) mice, it was observed that LCS enhanced the intestinal absorption of cholecalciferol by modulating vitamin D transporter protein levels and elevated 25OHD3 levels in senile osteoporosis (SOP). In Caco-2 cells, the addition of LCS significantly increased cholecalciferol uptake at medium concentrations of 0.5 μM (P = 0.007) and 10 μM (P = 0.025).27 Administration of 2.9 × 109 CFU of Limosilactobacillus reuteri NCIMB 30242 significantly increased serum 25-hydroxyvitamin D levels by 14.9 nmol L−1 (25.5%) in healthy hypercholesterolemic adults during the intervention period. This change represented a mean difference of 17.1 nmol L−1 (22.4%) compared to the placebo group, with statistical significance (P = 0.003).28 Mice administered probiotic cells received a daily oral gavage of 108 CFU of Lacticaseibacillus paracasei DG in combination with a vitamin D3 supplement for one week. Serum 25-hydroxyvitamin D [25(OH)D] levels in these mice ranged from 84 to 89 ng mL−1 (mean: 88 ± 2 ng mL−1), reflecting average increases of 50%, 62%, and 55% compared to the control group, the vitamin D3-only group, and the DG cells-alone group, respectively.29 Daily supplementation with compound probiotics (4.2 × 107 CFU g−1) containing Lactobacillus and Bifidobacterium for 10 weeks at a feeding rate equivalent to 1%–2% of total body weight significantly increased the activities of amylase (AMS), lipase (LPS), and trypsin (TPS), thereby improving the digestion and absorption of dietary vitamins.30 Probiotics enhance vitamin absorption, optimizing nutritional status, disease prevention, and overall health.
3. Factors affecting the intestinal absorption of vitamins promoted by probiotics
3.1. Intestinal microbiota structure
The importance of the gut microbiota in intestinal vitamin absorption cannot be overstated. The gut microbiota plays a critical role in facilitating vitamin absorption. Probiotics help alleviate intestinal inflammation and improve digestive function. They achieve this by normalizing the composition of the gut microbiota, preventing the colonization of pathogens in the digestive tract, enhancing intestinal barrier function, and modulating immune responses. This mechanism aids in the treatment or prevention of intestinal diseases such as inflammatory bowel disease (IBD), ulcerative colitis (UC), and Crohn's disease (CD), which are associated with impaired vitamin absorption (Fig. 2).31 | ||
| Fig. 2 Factors in probiotics that promote the absorption of vitamins in the gut. E. coli: Escherichia coli, H. pylori: Helicobacter pylori, E. faecalis: Enterococcus faecalis. | ||
A recent analysis confirmed the negative impact of chronic inflammation and intestinal injury on intestinal absorption of vitamin D. The results showed that mean vitamin D levels were 3.34 ng mL−1 (8.36 nmol L−1) lower in CD patients than in controls.32 Inflammation and Helicobacter pylori infection can cause malabsorption of dietary vitamin B12.33 Additionally, probiotic supplementation has been found to mitigate several gastrointestinal disorders, including IBD and irritable bowel syndrome (IBS), and increase the eradication rate of Helicobacter pylori.34 Some species of Lactobacillus, such as Lactobacillus acidophilus, Lactobacillus bulgaricus, Lacticaseibacillus casei, and Lacticaseibacillus rhamnosus GG, have been shown to prevent antibiotic-associated diarrhea and Clostridium difficile-associated colitis while maintaining the physiological microecology of the intestinal tract and providing an environment conducive to vitamin absorption.35Lacticaseibacillus paracasei LC-37 relieved functional dyspepsia (FD) in participants after 14 and 28 days of treatment with an LC-37-containing beverage. Lacticaseibacillus paracasei LC-37 significantly reduced clinical symptom scores, with abdominal pain and belching markedly decreasing after 14 days and nearly disappearing by 28 days of treatment. Lacticaseibacillus paracasei LC-37 increased the abundance of beneficial probiotics such as Lactobacillus, Lactococcus, and Weissella, while significantly reducing harmful bacteria like Lachnoclostridium.36 Numerous studies have emphasized the importance of bifidobacteria in the regulation of intestinal homeostasis, modulation of local and systemic immune responses, and the prevention of inflammatory diseases and infections. Bifidobacteria can convert certain dietary compounds into vitamins and specific strains have been shown to promote the absorption of vitamins in the intestine.37 The gut microbiota plays a critical role in vitamin absorption, which may be adversely affected by changes in composition and diversity. Probiotic supplementation, however, can improve vitamin absorption and maintain gut health. Overall, the strategic use of probiotics can significantly enhance the intestinal absorption of vitamins, thereby contributing to improved health outcomes and disease prevention.
3.2. Intestinal pH
The acid–base environment of the intestine is an important factor that influences the intestinal absorption of vitamins, which may have important effects on food solubility, vitamin release, vitamin stability, and intestinal permeability. An acidic pH-dependent mechanism influences intestinal absorption of vitamin B3.38 Specifically, when the extracellular pH of Caco-2 cells decreased to 5.0, the cellular uptake of vitamin B3 increased. When vitamin D3 is absorbed through a passive diffusion mechanism in the intestine, its absorption rate increases as pH decreases.39 Probiotics can decrease the intestinal pH by producing organic acids, which can increase vitamin absorption. Butyricicoccus pullicaecorum is a bacterium with probiotic potential that produces butyrate, which has a positive effect on gut health.40 Additionally, Limosilactobacillus reuteri, with its BSH activity, may decrease intestinal pH and increase vitamin D absorption and utilization by producing lactate.28 Lactic acid bacteria, such as Bifidobacterium, Lactobacillus, Enterococcus, and Streptococcus, in the intestinal tract produce lactic acid, which reduces local acidity in the intestinal lumen and promotes the absorption of most B vitamins and vitamin C.7 Acidic conditions in the intestine protect vitamin C from chemical and enzymatic oxidation. However, increased intestinal pH can cause oxidation of vitamin C or complex formation with other components, leading to decreased intestinal absorption of vitamin C.41 Probiotics can lower the intestinal pH by producing organic acids and enhancing vitamin solubility, release, stability, and intestinal permeability, thereby increasing vitamin absorption. Therefore, probiotic supplementation can optimize the intestinal environment and maintain intestinal pH balance, which are essential for improving the efficiency of vitamin absorption.3.3. Enzyme activity in the gut
Dietary vitamins are absorbed through the release of free-form vitamins from dietary proteins, which are bound to them in the presence of enzymes. These vitamins must first be transported to enterocytes and the intestine via specific carriers. When gastric lipase activity is reduced or insufficient, fat-soluble vitamin deficiency can occur. For example, vitamin A requires pancreatic carboxyl ester lipase (CEL) to break it down, and vitamin E is passively absorbed by intestinal epithelial cells in the form of mixed micelles after lipase action. CEL knockout (CELKO) mice can only absorb approximately 50% of the dietary vitamin A compared to wild type (WT) mice.42 Similarly, water-soluble vitamins require specific enzymatic action for intestinal absorption. For instance, vitamin B12 conjugates with proteins, which requires the action of proteases to release free vitamin B12. Subsequently, in the proximal intestine, trypsin degrades cryptoglobulin, allowing free vitamin B12 to bind to intrinsic factors secreted by gastric parietal cells.43 In addition, a decrease in the activity of vitamin B12-dependent methionine synthase results in a proportional deficiency in vitamin B12.Probiotics can promote the digestion and absorption of vitamins by secreting or enhancing the enzymatic activity of key digestive enzymes such as protease, lipase, amylase, and cellulase, which are present in the intestine. Previous studies have demonstrated that probiotics can increase the activity of antioxidant enzymes in aquatic animals, which attenuates oxidative stress, allowing epithelial cells of the small intestine to maintain their vitamin absorptive function.44 Dual-species probiotics, specifically Bacillus subtilis and Lactiplantibacillus plantarum, have been shown to significantly improve digestive enzyme activities, including total protease, α-amylase, lipase, and trypsin, as well as intestinal antioxidant capacity, as measured by superoxide dismutase, malondialdehyde, and catalase, in grass carp45 and white-spotted moray eel. Probiotics play an important role in the absorption of vitamins by increasing the activity of digestive enzymes, which further facilitates the maintenance of the nutritional status of the vitamins released from food and improves intestinal absorption.
3.4. Intestinal peristalsis
Although the enteric nervous system (ENS) regulates intestinal physiological processes, it is essential to acknowledge the impact of the intestinal environment on the physiology of the intestinal tissues.46 In general, after eating, the muscles in the intestinal and gastric walls of individuals hold slow, regular peristalsis for several hours and secrete digestive juices, enzymes, and choline, which mix with digested food in contact with the intestinal lining.47 During this process, the vitamins are absorbed by the intestinal tract. Without peristaltic movement, there would be no digestion or absorption of vitamins from the food. Numerous studies have confirmed that probiotics can alter intestinal activity and affect intestinal peristalsis and secretion, thus benefiting patients with intestinal dyspareunia.48,49 An experiment using zebrafish to screen probiotics for intestinal peristalsis rapidly revealed that all the selected probiotics improved intestinal peristalsis. Different strains exhibit different effects on intestinal peristalsis. Lacticaseibacillus paracasei X11 effectively promotes intestinal peristalsis.50Lactobacillus royale and Lactobacillus gattii OLL2716 can slow gastric emptying, potentially affecting vitamin B12 absorption. Daily probiotic treatment with Lactobacillus brevis is highly beneficial for vitamin B12 absorption.7 Supplementation with products containing Lactobacillus or Bifidobacterium species increases intestinal motility and decreases intestinal transit time (ITT) in adults.51 Probiotics can help improve intestinal dysbiosis by altering intestinal activity, motility, and secretion and promoting vitamin absorption, particularly in cases of abnormal motility.3.5. Gastrointestinal transit and binding proteins
Probiotics have been demonstrated to affect the synthesis of vitamin transporter proteins, which subsequently promote the intestinal absorption of vitamins. Lacticaseibacillus rhamnosus GG and Lactiplantibacillus plantarum8 can influence the plasma membrane of the intestinal cells to promote the synthesis of NPC1L1, a key transporter protein for lipids.58,59 When mice were administered 1 ml day−1 of Lacticaseibacillus rhamnosus GG culture supernatant (LCS) for four weeks, a significant increase in NPC1L1 and CD36 and a decrease in SR-BI were observed in the proximal small intestine of mice. This resulted in the enhanced intestinal absorption of vitamin D.27 Intestinal absorption of fat-soluble vitamins, such as vitamin E and vitamin A (e.g., β-carotene and lutein), enters the intestinal epithelium mainly through the SR-BI transporter protein.60 However, the absorption of these vitamins requires lipid involvement.20 Some probiotics reduce lipid absorption by inhibiting the expression of lipid transporter proteins,8 which indirectly leads to reduced intestinal absorption of fat-soluble vitamins, such as carotenoids, vitamin E and vitamin K. Probiotics influence the synthesis of transporter proteins that are essential for their selective and specific absorption. This process ultimately facilitates the intestinal absorption of vitamins, especially fat-soluble vitamins.
3.6. Dietary lipids and cholesterol
The co-consumption of dietary lipids promotes the absorption of fat-soluble vitamins and disruption of lipid absorption leads to reduced absorption of fat-soluble vitamins.28 Cholesterol, which is structurally homologous to vitamin D, competes with vitamin D for intestinal absorption. Probiotics such as Bifidobacterium and Lactobacillus can affect the intestinal absorption of fat-soluble vitamins through direct and indirect involvement in lipid metabolism. For example, Lactobacillus acidophilus can promote vitamin D absorption by eliminating cholesterol69 and producing lipids, such as short-chain fatty acids (SCFAs), which increase the bioavailability of vitamins, making them more soluble and absorbable.70 Probiotics such as Enterococcus faecalis, Lactobacillus pentosus, and Lactobacillus possess the dual potential of producing vitamin B12 and lowering cholesterol. This makes them a valuable source of vitamin B12 and help in breaking down excess lipid molecules, thereby promoting the absorption of fat-soluble vitamins and vitamin B12. They have been used to overcome vitamin B12 deficiency and treat hypercholesterolemia.71 Overall, probiotics help overcome vitamin B12 deficiency by participating in lipid metabolism, influencing cholesterol removal, producing lipids, such as short-chain fatty acids, promoting intestinal absorption of fat-soluble vitamins, and acting as a reliable source of vitamin B12.4. Possible mechanisms of action of probiotics on intestinal vitamin absorption
4.1. Probiotics change the composition of the intestinal microbiota
Probiotics, such as Lactobacillus and Bifidobacterium, affect the gut–brain axis, gut–kidney axis, and gut–liver, competing with pathogens for nutrients and binding sites in the gut wall and affecting the composition of the intestinal microbiota. They modulate biosynthetic processes and absorption and directly or indirectly improve vitamin absorption.6,72–74 Probiotics produce metabolites, including bacterial components and postbiotics,75 which inhibit the growth of harmful bacteria and improve the structure and function of the intestinal microbiota,76 thereby providing specific physiological effects that improve host health and favor vitamin absorption (Fig. 3). | ||
| Fig. 3 Probiotics promote vitamin absorption by altering the composition of the intestinal microbiota. Slc19a2: solute carrier family 19 Member 2, Slc19a3: solute carrier family 19 Member 3. | ||
Probiotics can interact with the gut microbiota to compete for nutrients, display antagonism, engage in cross-feeding, and support microbiota stability. This can optimize the composition of the gut microbiota and promote vitamin absorption by the gut.77
Gram-negative bacteria such as Escherichia coli, which contain a significant portion of the intestinal microbiota, release substantial amounts of lipopolysaccharides to impede the function of solute carrier family 23 member 1 (SLC23A1) and solute carrier family 23 member 2 (SLC23A2) promoters. Consequently, the expression of sodium-dependent vitamin C transport proteins SVCT1 and SVCT2 is reduced, leading to decreased vitamin C absorption.78 In contrast, enterotoxin-producing Escherichia coli diminishes the expression levels of THTR-1 and THTR-2, resulting in a significant reduction in vitamin B1 absorption.79Lacticaseibacillus rhamnosus GG has been shown to mitigate the impairment of the intestinal barrier function by Escherichia coli by fine-tuning the composition of the intestinal microbiota, specifically by increasing the abundance of dead Escherichia coli.80,81 In addition to its capacity for vitamin B9 biosynthesis, Lactiplantibacillus plantarum demonstrates a distinctive modulatory effect on the gut microbiota composition through a multi-stage cascade mechanism. This probiotic strain induces sequential enrichment of specific bacterial taxa, including Adlercreutzia → Akkermansia, p_75_a5, Coprococcus → Allobaculum, Dehalobacterium, Dorea, Parabacteroides. Through this cascading network of microbial interactions, L. plantarum effectively reconstitutes the compromised architecture of intestinal microbiota communities. Such structural restoration establishes an optimized microenvironment that enhances the intestinal absorption efficiency of vitamin B9.82,83 The combined use of Limosilactobacillus fermentum HF06 and Lactiplatibacillus plantarum HF05 has been shown to modulate the structure and composition of the intestinal microbiota and promote the abundance of beneficial bacteria to decrease the abundance of pro-inflammatory and liver injury-associated bacteria, such as Clostridium Sensu Stricto, Escherichia_Shigella, Enterococcus, Corynebacterium, Desulfovibrio and Norank-f-Oscillospiraceae, effectively reduce intestinal inflammation, and improve vitamin absorption in patients.84
4.2. Probiotics regulate intestinal pH
Vitamins are highly susceptible to oxidation and inactivation in neutral and alkaline environments. However, probiotics possess high acidifying activity, producing a significant amount of organic acids such as lactic acid, acetic acid, and propionic acid during the breakdown of food. This results in a lower pH in the gut, which helps to maintain vitamin activity. Probiotics also regulate intestinal pH and influence the activity of digestive enzymes, thereby improving intestinal digestion and increasing vitamin absorption.85 In addition, a lower pH intestinal environment inhibits the growth of pathogenic bacteria, such as Staphylococcus aureus and Escherichia coli. In contrast, probiotic bacteria that are tolerant to acidic conditions can survive in the lower pH environment of the intestines.86Certain bacteria, such as Bifidobacterium, Lactobacillus, Lacticaseibacillus casei GG,16Enterococcus, and Streptococcus, rapidly metabolize carbohydrates into organic acids, including acetic and lactic acids, which regulate intestinal pH and create an acidic environment that is unfavorable for pathogen growth and enhances the absorption of vitamin B. Lactobacillus and Bifidobacterium produce lactic acid and acetic acid as major end products of carbohydrate metabolism. When produced in situ, these organic acids reduce the intracavitary pH and inhibit the growth of pathogenic bacteria.87,88 Bifidobacterium primarily produces SCFAs through fermentation. In a simulated colon environment, the oligosaccharides from Chinese yams can serve as carbon sources for intestinal probiotics such as Lactobacillus plantarum and Bifidobacterium. After 48 hours of fermentation, the acetic acid content in the Bifidobacterium culture reached 1.85 mg mL−1, whereas, after 8 hours, it was 0.082 mg mL−1.89 SCFAs are produced through multiple metabolic pathways. The most common pathway is glycolysis, while certain bacterial groups, such as Bifidobacterium, can also utilize the pentose phosphate pathway to generate these metabolites.90
4.3. Probiotics regulate intestinal metabolism and function
Intestinal metabolism and function are closely related to the intestinal environment, and changes in the intestinal environment have important implications for vitamin absorption. Probiotics have been found to enhance intestinal function, improve the intestinal mucosal barrier function, and regulate immune function. The production of active substances and enzyme formation promote vitamin absorption in several ways (Fig. 4).77,91–94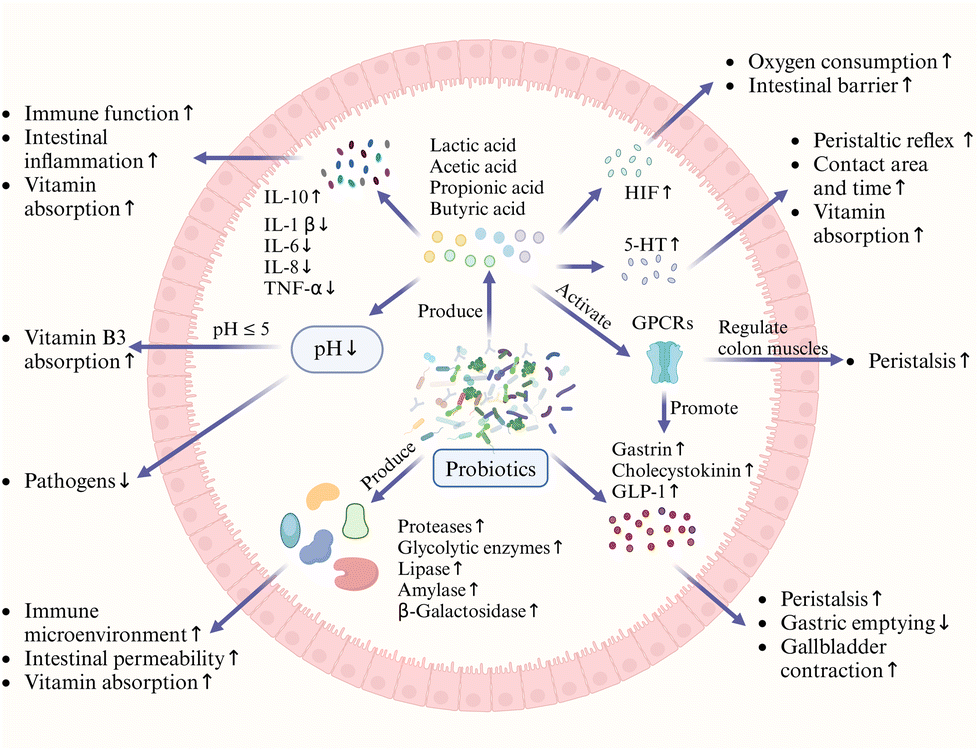 | ||
| Fig. 4 Probiotics modulate gut metabolism and vitamin absorption. GPCRs: g protein-coupled receptors. | ||
The metabolism of probiotics can produce butyric acid, which promotes the consumption of oxygen by intestinal epithelial cells, thereby increasing the expression of barrier-protective hypoxia-inducible factor (HIF) target genes and maintaining the stability of HIF.95 Probiotic metabolism produces butyrate, propionate, and acetate, which are involved in evolutionarily conserved processes such as modulation of epithelial barrier function and mucosal and systemic immunity. These effects are mediated through various mechanisms, including G-protein-coupled receptor signaling and histone deacetylase activity. Additionally, the anti-inflammatory effects of butyrate are mediated through direct effects on various cell types, including intestinal epithelial cells, phagocytes, B cells, plasma cells, and regulatory and effector T cells.74,96 These mechanisms include the initiation of pattern recognition receptors and changes in bidirectional connectivity between the gastrointestinal tract (GIT) and the central nervous system (CNS).97 Furthermore, probiotics can affect gut hormones such as gastrin, cholecystokinin, and glucagon-like peptide-1 (GLP-1), which play important roles in controlling bowel motility. For example, GLP-1 slows gastric emptying, allowing the gut to absorb nutrients for more time. Lipid postbiotics produced by probiotics also delay gastric emptying and nutrient transfer through the small intestine, providing additional time for fat-soluble vitamins to be released into the micelle mixture and dissolve, thereby facilitating their intestinal uptake.98,99 Probiotics also produce a variety of vitamins and enzymes, such as proteases, glycolytic enzymes, lipases, amylases, and β-galactosidase, which improve the immune microenvironment, enhance intestinal permeability, and facilitate the intestinal digestion and absorption of vitamins.100 Specifically, Bifidobacterium spp., Lactobacillus spp., and Streptococcus spp. can stimulate ileal segmental contraction, enhance intestinal peristalsis by using SCFAs to directly act on the intestinal smooth muscle or through the activation of intestinal receptors (e.g., G-protein-coupled receptors), and promote the release of intestinal hormones such as ghrelin and cholecystokinin. These hormones help regulate the contraction and relaxation of colonic muscles, thereby playing an active role in regulating intestinal motility and promoting vitamin absorption.101Lactiplantibacillus plantarum BMCM12 has been shown to secrete extracellular proteins that can attenuate pathogen adhesion and protect the intestinal barrier.
Ingestion of Lactobacillus has been demonstrated to contribute to the maintenance of epithelial permeability, enhancement of the mucosal layer, increase of intestinal cell turnover, and stimulation of both innate and adaptive immune responses.102 When intestinal disease occurs, enteric nerve activation of the Ahr gene, which enables normal peristalsis of the colon, may be disrupted,46 which results in the accumulation of significant amounts of SCFAs (mainly acetate and propionate and butyrate), which are released through the action of 5-hydroxytryptamine (5-HT) to initiate or enhance the peristaltic reflex. This increases the area and duration of contact between the vitamin and the intestine, thereby promoting its absorption.103,104 Patients with IBD have significantly lower levels of vitamin absorption than the normal population, which puts them at a risk of developing vitamin deficiencies.105,106 Numerous species within the genus Lactobacillus have been identified as having potential therapeutic benefits in the treatment of IBD.107 Short-chain fatty acids, especially butyric acid, are known to stimulate immune function in animal and human cells while inhibiting the expression of pro-inflammatory cytokines interleukin-1 β (IL-1 β), interleukin-6 (IL-6), interleukin-8 (IL-8), and tumor necrosis factor-α (TNF-α), and inducing the expression of the anti-inflammatory cytokine interleukin-10 (IL-10). They improve intestinal inflammation and develop favorable conditions for vitamin absorption.108Lactiplantibacillus plantarum, an immunomodulatory probiotic, modulates the immune response of gut-associated lymphocytes and epithelial cells by colonizing the gut after a GIT transport process. This stimulates the production of immune factors that regulate the ratio of T helper 1 (Th1) to T helper 2 (Th2) cells, thus alleviating inflammatory bowel disease.109 In contrast, fermentation of Limosilactobacillus fermentum results in markers involved in the immune response, as well as the expression of microRNA-155 (miR-155) and microRNA-223 (miR-223). Furthermore, it improved the alterations in the expression of microRNA-150 (miR-150) and microRNA-143 (miR-143), which are involved in the maintenance of the barrier function. Additionally, it inhibits the increase in the expression of interleukin-12 (IL-12) and inducible nitric oxide synthase (iNOS) and recovers vitamin absorption.110 Overall, the ability of probiotics to modulate the gut microbiota and produce beneficial metabolites underscores their critical role in regulating intestinal metabolism and function, providing a promising avenue for therapeutic interventions in various metabolic and gastrointestinal disorders.
4.4. Probiotics affect transporter proteins
Vitamin uptake from the lumen to the cytoplasmic lysate of enterocytes involves specific membrane transport proteins.38,111 Researchers examined the genomic findings of transporter proteins in various probiotic bacteria and found that all ten Lactobacillus strains possess a significant number of vitamin transporters that facilitate intestinal vitamin absorption.112 However, no studies have elucidated the precise mechanisms by which probiotics affect the synthesis of vitamin transporter proteins (Fig. 5). Therefore, further specialized in-depth studies are required. | ||
| Fig. 5 Mechanisms probiotics use to influence transporter proteins to promote intestinal absorption of vitamin D. | ||
Current evidence indicates that probiotics influence vitamin transporter synthesis through the regulation of gene expression associated with nutrient transport, thereby modulating intestinal vitamin absorption. A multi-species probiotic (MSP) comprising Bacillus coagulans, Bacillus subtilis, and Saccharomyces boulardii has demonstrated a significant effect in enhancing the expression of vitamin transporter genes, such as sodium-dependent glucose transporter (SGLT1) in chicks.113 Similarly, a combination of Lactobacillus acidophilus, Bacillus subtilis, and Enterococcus faecalis has been shown to upregulate nutrient transport-related genes, including sodium–glucose co-transporter-1 (SGLT-1), glucose transporter-2 (GLUT-2), cationic amino acid transporter-1 (CAT-1), and fatty acid-binding protein-2 (FABP-2), in rabbits, while also altering cytokine expression and ultimately promoting intestinal vitamin absorption.114Lactobacillus acidophilus has been shown to contribute to the reduction of NPCIL1 by decreasing NPCIL1 gene expression and inhibiting the uptake of micellar cholesterol and fat-soluble vitamins in Caco-2 cells, and potentially mediating this effect through the liver X receptor (LXR).115 Overall, probiotics play diverse and significant roles in modulating transporter proteins, which can have therapeutic implications in various health conditions, including gastrointestinal diseases, metabolic disorders, and cancer.
5. Conclusion and prospects
This review systematically delineates the absorption mechanisms of fat-soluble and water-soluble vitamins in the gut (active transport at low concentrations and passive diffusion at high concentrations). It elucidates the regulatory roles of host intestinal microenvironments (e.g., pH, enzymatic activity, and microbiota composition) in vitamin absorption. It highlights how probiotics directly or indirectly modulate vitamin transporter expression and lipid absorption efficiency through pathways including organic acid secretion to regulate intestinal pH, enhancement of digestive/antioxidant enzyme activity, competitive inhibition of pathogenic colonization, immunomodulation, and microbiota remodeling via metabolites (e.g., postbiotics). Furthermore, probiotics synergistically enhance vitamin absorption by strengthening mucosal barrier function, regulating systemic metabolism, and balancing intestinal cell proliferation/apoptosis through programmed regulation. These multidimensional mechanisms provide a theoretical foundation and innovative nutritional intervention strategy for addressing vitamin deficiencies through probiotic-based alternatives to conventional vitamin fortification approaches.Significant knowledge gaps remain in elucidating the mechanisms by which probiotics regulate intestinal vitamin absorption and their clinical translation applications. Current evidence suggests that probiotics primarily influence vitamin metabolism through the following pathways: modulating gut microenvironment homeostasis, secreting bioactive metabolites, and participating in host–microbiota interaction networks. However, the specific molecular targets, strain-specific functions, and host-dependent response mechanisms have not yet been fully clarified. Notably, precision intervention strategies based on gut microbiota profiles and host genetic polymorphisms remain in the exploratory stage. Furthermore, long-term safety assessment frameworks and evidence-based clinical translation pathways still require refinement.
Future research should focus on the following four aspects to deepen the understanding of the synergistic effects between probiotics and vitamins and provide practical guidance for clinical applications: (a) adopt multi-omics integrated analysis strategies to dynamically dissect the spatiotemporal remodeling features of vitamin metabolic networks during probiotic colonization, with an emphasis on elucidating the molecular interaction mechanisms between microbiota-derived metabolites and host vitamin transporters; (b) screen probiotic strains with high-efficiency vitamin synthesis capabilities and investigate the additive effects of strain combinations (symbiotic or synergistic interactions) on multi-dimensional nutrient absorption; (c) develop personalized probiotic formulations based on individual gut microbiota profiles and metabolic phenotypes; and (d) conduct large-scale randomized controlled trials (RCTs) to clarify the adjunctive therapeutic value of specific probiotics in addressing vitamin deficiency disorders.
Author contributions
Peiling Hu: conceptualization, methodology, formal analysis, investigation, writing – original draft, and visualization; Han Tan: formal analysis and visualization; Weiping Xu: formal analysis and visualization; Shijian Liu: supervision; Jiajia Song: supervision; Xiaoyong Chen: conceptualization, methodology, and writing – reviewing and editing; Huayi Suo: project administration, funding acquisition, and supervision. All authors wrote the manuscript and approved its final version.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This work was financially supported by the Sichuan Province International Science and Technology Innovation Cooperation/Hong Kong, Macao and Taiwan Science and Technology Innovation Cooperation project (Grant No. 2023YFH0002). We want to express our gratitude for the drawing materials provided by BioRender.References
- J. Timoneda, L. Rodríguez-Fernández, R. Zaragozá, M. Marín, M. Cabezuelo, L. Torres, J. Viña and T. Barber, Vitamin A Deficiency and the Lung, Nutrients, 2018, 10, 1132 CrossRef PubMed.
- G. Yu, S. Liu, K. Yang and Q. Wu, Reproductive-dependent effects of B vitamin deficiency on lifespan and physiology, Front. Nutr., 2023, 10, 1277715 CrossRef PubMed.
- M. Nguyen, The Forgotten Story of Vitamin C Deficiency, Blood, 2020, 136, 35 CrossRef.
- M. F. Holick, Vitamin D: Important for Prevention of Osteoporosis, Cardiovascular Heart Disease, Type 1 Diabetes, Autoimmune Diseases, and Some Cancers, South. Med. J., 2005, 98, 1024–1026 CrossRef PubMed.
- C. Hill, F. Guarner, G. Reid, G. R. Gibson, D. J. Merenstein, B. Pot, L. Morelli, R. B. Canani, H. J. Flint, S. Salminen, P. C. Calder and M. E. Sanders, The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic, Nat. Rev. Gastroenterol. Hepatol., 2014, 11, 506–514 Search PubMed.
- C. Hu, L. Tang, M. Liu, P. K. S. Lam, J. C. W. Lam and L. Chen, Probiotic modulation of perfluorobutanesulfonate toxicity in zebrafish: Disturbances in retinoid metabolism and visual physiology, Chemosphere, 2020, 258, 127409 CrossRef CAS PubMed.
- Z. Wan, J. Zheng, Z. Zhu, L. Sang, J. Zhu, S. Luo, Y. Zhao, R. Wang, Y. Zhang, K. Hao, L. Chen, J. Du, J. Kan and H. He, Intermediate role of gut microbiota in vitamin B nutrition and its influences on human health, Front. Nutr., 2022, 9, 1031502 CrossRef PubMed.
- H.-S. Yoon, J.-H. Ju, H.-N. Kim, H.-J. Park, Y. Ji, J.-E. Lee, H.-K. Shin, M.-S. Do and W. Holzapfel, Reduction in cholesterol absorption in Caco-2 cells through the down-regulation of Niemann-Pick C1-like 1 by the putative probiotic strains Lactobacillus rhamnosus BFE5264 and Lactobacillus plantarum NR74 from fermented foods, Int. J. Food Sci. Nutr., 2013, 64, 44–52 Search PubMed.
- S. Magnúsdóttir, D. Ravcheev, V. De Crécy-Lagard and I. Thiele, Systematic genome assessment of B-vitamin biosynthesis suggests co-operation among gut microbes, Front. Genet., 2015, 6, 148 Search PubMed.
- P. Eck, Nutrigenomics of vitamin C absorption and transport, Curr. Opin. Food Sci., 2018, 20, 100–104 CrossRef.
- R. Zhao and I. D. Goldman, Folate and thiamine transporters mediated by facilitative carriers (SLC19A1–3 and SLC46A1) and folate receptors, Mol. Aspects Med., 2013, 34, 373–385 CrossRef CAS PubMed.
- A. Yonezawa and K. Inui, Novel riboflavin transporter family RFVT/SLC52: Identification, nomenclature, functional characterization and genetic diseases of RFVT/SLC52, Mol. Aspects Med., 2013, 34, 693–701 CrossRef CAS PubMed.
- D. Kibirige and R. Mwebaze, Vitamin B12 deficiency among patients with diabetes mellitus: is routine screening and supplementation justified?, J. Diabetes Metab. Disord., 2013, 12, 17 CrossRef PubMed.
- H. M. Said, A. Ortiz and T. Y. Ma, A carrier-mediated mechanism for pyridoxine uptake by human intestinal epithelial Caco-2 cells: regulation by a PKA-mediated pathway, Am. J. Physiol.: Cell Physiol., 2003, 285, C1219–C1225 CrossRef CAS PubMed.
- A. Brito, E. Habeych, I. Silva-Zolezzi, N. Galaffu and L. H. Allen, Methods to assess vitamin B12 bioavailability and technologies to enhance its absorption, Nutr. Rev., 2018, 76, 778–792 CrossRef PubMed.
- I. Elmadfa, C. Heinzle, D. Majchrzak and H. Foissy, Influence of a Probiotic Yoghurt on the Status of Vitamins B1, B2 and B6 in the Healthy Adult Human, Ann. Nutr. Metab., 2001, 45, 13–18 CrossRef CAS PubMed.
- E. Fabian, D. Majchrzak, B. Dieminger, E. Meyer and I. Elmadfa, Influence of Probiotic and Conventional Yoghurt on the Status of Vitamins B1, B2 and B6 in Young Healthy Women, Ann. Nutr. Metab., 2008, 52, 29–36 CrossRef CAS PubMed.
- A. Pompei, L. Cordisco, A. Amaretti, S. Zanoni, S. Raimondi, D. Matteuzzi and M. Rossi, Administration of Folate-Producing Bifidobacteria Enhances Folate Status in Wistar Rats, J. Nutr., 2007, 137, 2742–2746 CrossRef CAS PubMed.
- V. C. Molina, M. Médici, M. P. Taranto and G. Font De Valdez, Lactobacillus reuteri CRL 1098 prevents side effects produced by a nutritional vitamin B 12 deficiency, J. Appl. Microbiol., 2009, 106, 467–473 CrossRef CAS PubMed.
- P. Borel, O. Dangles and R. E. Kopec, Fat-soluble vitamin and phytochemical metabolites: Production, gastrointestinal absorption, and health effects, Prog. Lipid Res., 2023, 90, 101220 CrossRef CAS PubMed.
- D. Hollander, K. S. Muralidhara and A. Zimmerman, Vitamin D-3 intestinal absorption in vivo: influence of fatty acids, bile salts, and perfusate pH on absorption, Gut, 1978, 19, 267–272 CrossRef CAS PubMed.
- P. Borel, Factors Affecting Intestinal Absorption of Highly Lipophilic Food Microconstituents (Fat-Soluble Vitamins, Carotenoids and Phytosterols), Clin. Chem. Lab. Med., 2003, 41, 979–994 CAS.
- A. Van Bennekum, M. Werder, S. T. Thuahnai, C.-H. Han, P. Duong, D. L. Williams, P. Wettstein, G. Schulthess, M. C. Phillips and H. Hauser, Class B Scavenger Receptor-Mediated Intestinal Absorption of Dietary β-Carotene and Cholesterol, Biochemistry, 2005, 44, 4517–4525 CrossRef CAS PubMed.
- E. Reboul, A. Goncalves, C. Comera, R. Bott, M. Nowicki, J. Landrier, D. Jourdheuil-Rahmani, C. Dufour, X. Collet and P. Borel, Vitamin D intestinal absorption is not a simple passive diffusion: Evidences for involvement of cholesterol transporters, Mol. Nutr. Food Res., 2011, 55, 691–702 CrossRef CAS PubMed.
- Y. Yamanashi, T. Takada, R. Kurauchi, Y. Tanaka, T. Komine and H. Suzuki, Transporters for the Intestinal Absorption of Cholesterol, Vitamin E, and Vitamin K, J. Atheroscler. Thromb., 2017, 24, 347–359 CrossRef CAS PubMed.
- A. Goncalves, M. Margier, S. Roi, X. Collet, I. Niot, P. Goupy, C. Caris-Veyrat and E. Reboul, Intestinal Scavenger Receptors Are Involved in Vitamin K1 Absorption, J. Biol. Chem., 2014, 289, 30743–30752 CrossRef CAS PubMed.
- J. Cheng, J. Zhai, W. Zhong, J. Zhao, L. Zhou and B. Wang, Lactobacillus rhamnosus GG Promotes Intestinal Vitamin D Absorption by Upregulating Vitamin D Transporters in Senile Osteoporosis, Calcif. Tissue Int., 2022, 111, 162–170 CrossRef CAS PubMed.
- M. L. Jones, C. J. Martoni and S. Prakash, Oral Supplementation With Probiotic L. reuteri NCIMB 30242 Increases Mean Circulating 25-Hydroxyvitamin D: A Post Hoc Analysis of a Randomized Controlled Trial, J. Clin. Endocrinol. Metab., 2013, 98, 2944–2951 CrossRef CAS PubMed.
- I. Castagliuolo, M. Scarpa, P. Brun, G. Bernabe, V. Sagheddu, M. Elli, W. Fiore, V. De Vitis and S. Guglielmetti, Co-administration of vitamin D3 and Lacticaseibacillus paracasei DG increase 25-hydroxyvitamin D serum levels in mice, Ann. Microbiol., 2021, 71, 42 CrossRef CAS PubMed.
- X. Zhao, B. Shi, C. Wang, H. Cheng, C. Wang, X. Ma, K. Yan and Y. Chen, Effects of dietary supplementation of compound probiotics on growth, non-specific immunity, intestinal digestive enzyme, and microbiota of whitespotted conger (conger myriaster) in recirculating aquaculture system, Aquacult. Int., 2024, 32, 2713–2738 CrossRef CAS.
- M. Mikelsaar and M. Zilmer, Lactobacillus fermentum ME-3 – an antimicrobial and antioxidative probiotic, Microb. Ecol. Health Dis., 2009, 21, 1–27 CrossRef CAS PubMed.
- A. Giustina, L. di Filippo, A. Allora, D. D. Bikle, G. M. Cavestro, D. Feldman, G. Latella, S. Minisola, N. Napoli, S. Trasciatti, M. Uygur and J. P. Bilezikian, Rev. Endocr. Vitamin D and malabsorptive gastrointestinal conditions: A bidirectional relationship?, Rev. Endocr. Metab. Disord., 2023, 24, 121–138 CrossRef CAS PubMed.
- R. Kozyraki and O. Cases, Vitamin B12 absorption: Mammalian physiology and acquired and inherited disorders, Biochimie, 2013, 95, 1002–1007 CrossRef CAS PubMed.
- J. L. Tong, Z. H. Ran, J. Shen, C. X. Zhang and S. D. Xiao, Meta–analysis: the effect of supplementation with probiotics on eradication rates and adverse events during Helicobacter pylori eradication therapy, Aliment. Pharmacol. Ther., 2007, 25, 155–168 CrossRef CAS PubMed.
- C. Iino, T. Shimoyama, D. Chinda, T. Arai, D. Chiba, S. Nakaji and S. Fukuda, Infection of Helicobacter pylori and Atrophic Gastritis Influence Lactobacillus in Gut Microbiota in a Japanese Population, Front. Immunol., 2018, 9, 712 CrossRef PubMed.
- E. Sun, X. Zhang, Y. Zhao, J. Li, J. Sun, Z. Mu and R. Wang, Beverages containing lactobacillus paracasei LC-37 improved functional dyspepsia through regulation of the intestinal microbiota and their metabolites, J. Dairy Sci., 2021, 104, 6389–6398 CrossRef CAS PubMed.
- J. G. LeBlanc, C. Milani, G. S. De Giori, F. Sesma, D. Van Sinderen and M. Ventura, Bacteria as vitamin suppliers to their host: a gut microbiota perspective, Curr. Opin. Biotechnol., 2013, 24, 160–168 CrossRef CAS PubMed.
- H. Takanaga, H. Maeda, H. Yabuuchi, I. Tamai, H. Higashida and A. Tsuji, Nicotinic Acid Transport Mediated by pH-dependent Anion Antiporter and Proton Cotransporter in Rabbit Intestinal Brush-border Membrane, J. Pharm. Pharmacol., 2011, 48, 1073–1077 CrossRef PubMed.
- D. Hollander, E. Rim and D. Morgan, Intestinal absorption of 25-hydroxyvitamin D3 in unanesthetized rat, Am. J. Physiol.: Endocrinol. Metab., 1979, 236, E441 Search PubMed.
- T. Gryp, G. R. B. Huys, M. Joossens, W. Van Biesen, G. Glorieux and M. Vaneechoutte, Isolation and Quantification of Uremic Toxin Precursor-Generating Gut Bacteria in Chronic Kidney Disease Patients, Int. J. Mol. Sci., 2020, 21, 1986 CrossRef CAS PubMed.
- M. J. Rodríguez-Roque, M. A. Rojas-Graü, P. Elez-Martínez and O. Martín-Belloso, Changes in Vitamin C, Phenolic, and Carotenoid Profiles Throughout in Vitro Gastrointestinal Digestion of a Blended Fruit Juice, J. Agric. Food Chem., 2013, 61, 1859–1867 CrossRef PubMed.
- W. Weng, L. Li, A. M. Van Bennekum, S. H. Potter, E. H. Harrison, W. S. Blaner, J. L. Breslow and E. A. Fisher, Intestinal Absorption of Dietary Cholesteryl Ester Is Decreased but Retinyl Ester Absorption Is Normal in Carboxyl Ester Lipase Knockout Mice, Biochemistry, 1999, 38, 4143–4149 CrossRef CAS PubMed.
- J. C. Fyfe, M. Madsen, P. Højrup, E. I. Christensen, S. M. Tanner, A. De La Chapelle, Q. He and S. K. Moestrup, The functional cobalamin (vitamin B12)–intrinsic factor receptor is a novel complex of cubilin and amnionless, Blood, 2004, 103, 1573–1579 CrossRef CAS PubMed.
- M. Roche, F. W. Kemp, A. Agrawal, A. Attanasio, P. V. S. V. Neti, R. W. Howell and R. P. Ferraris, Marked changes in endogenous antioxidant expression precede vitamin A-, C-, and E-protectable, radiation-induced reductions in small intestinal nutrient transport, Free Radicals Biol. Med., 2011, 50, 55–65 CrossRef CAS PubMed.
- N. Luo, L. Wang, Z. Wang, B. Xiao, N. Wang, X. Yu, D. Wu and Z. Song, Effects of dietary supplementation of duo-strain probiotics with post-spraying technology on growth performance, digestive enzyme, antioxidant capacity and intestinal microbiota of grass carp (Ctenopharyngodon idella), Aquacult. Rep., 2022, 26, 101301 Search PubMed.
- Y. Obata, Á. Castaño, S. Boeing, A. C. Bon-Frauches, C. Fung, T. Fallesen, M. G. De Agüero, B. Yilmaz, R. Lopes, A. Huseynova, S. Horswell, M. R. Maradana, W. Boesmans, P. Vanden Berghe, A. J. Murray, B. Stockinger, A. J. Macpherson and V. Pachnis, Neuronal programming by microbiota regulates intestinal physiology, Nature, 2020, 578, 284–289 CrossRef CAS PubMed.
- D. Borgmann, E. Ciglieri, N. Biglari, C. Brandt, A. L. Cremer, H. Backes, M. Tittgemeyer, F. T. Wunderlich, J. C. Brüning and H. Fenselau, Gut-brain communication by distinct sensory neurons differently controls feeding and glucose metabolism, Cell Metab., 2021, 33, 1466–1482 CrossRef CAS PubMed.
- T. Ma, X. Xue, H. Tian, X. Zhou, J. Wang, Z. Zhao, M. Wang, J. Song, R. Feng, L. Li, C. Jing and F. Tian, Effect of the gut microbiota and their metabolites on postoperative intestinal motility and its underlying mechanisms, J. Transl. Med., 2023, 21, 349 Search PubMed.
- E. Dimidi, S. Christodoulides, S. M. Scott and K. Whelan, Mechanisms of Action of Probiotics and the Gastrointestinal Microbiota on Gut Motility and Constipation, Adv. Nutr., 2017, 8, 484–494 CrossRef CAS PubMed.
- Y. Lu, J. Zhang, H. Yi, Z. Zhang and L. Zhang, Screening of intestinal peristalsis-promoting probiotics based on a zebrafish model, Food Funct., 2019, 10, 2075–2082 RSC.
- L. E. Miller, Effects of probiotic-containing products on stool frequency and intestinal transit in constipated adults: systematic review and meta-analysis of randomized controlled trials, Ann. Gastroenterol., 2017, 30, 629 Search PubMed.
- M. V. T. Lobo, L. Huerta, N. Ruiz-Velasco, E. Teixeiro, P. De La Cueva, A. Celdrán, A. Martín-Hidalgo, M. A. Vega and R. Bragado, Localization of the Lipid Receptors CD36 and CLA-1/SR-BI in the Human Gastrointestinal Tract: Towards the Identification of Receptors Mediating the Intestinal Absorption of Dietary Lipids, J. Histochem. Cytochem., 2001, 49, 1253–1260 CrossRef CAS PubMed.
- M. Bonnefond-Ortega, J. Goudable, C. Chambrier and C. Bétry, L′absorption intestinale des vitamines hydrosolubles et liposolubles en pratique clinique, Nutr. Clin. Metab., 2018, 32, 57–66 CrossRef.
- A. Goncalves, B. Gleize, S. Roi, M. Nowicki, A. Dhaussy, A. Huertas, M.-J. Amiot and E. Reboul, Fatty acids affect micellar properties and modulate vitamin D uptake and basolateral efflux in Caco-2 cells, J. Nutr. Biochem., 2013, 24, 1751–1757 CrossRef CAS PubMed.
- V. Ganapathy, S. B. Smith and P. D. Prasad, SLC19: the folate/thiamine transporter family, Pfluegers Arch., 2004, 447, 641–646 CrossRef CAS PubMed.
- S. M. Nabokina, K. Inoue, V. S. Subramanian, J. E. Valle, H. Yuasa and H. M. Said, Molecular Identification and Functional Characterization of the Human Colonic Thiamine Pyrophosphate Transporter, J. Biol. Chem., 2014, 289, 4405–4416 CrossRef CAS PubMed.
- A. Dutt Vadlapudi, R. Krishna Vadlapatla and A. K. Mitra, Sodium Dependent Multivitamin Transporter (SMVT): A Potential Target for Drug Delivery, Curr. Drug Targets, 2012, 13, 994–1003 CrossRef PubMed.
- C.-Y. Zhong, W.-W. Sun, Y. Ma, H. Zhu, P. Yang, H. Wei, B.-H. Zeng, Q. Zhang, Y. Liu, W.-X. Li, Y. Chen, L. Yu and Z.-Y. Song, Microbiota prevents cholesterol loss from the body by regulating host gene expression in mice, Sci. Rep., 2015, 5, 10512 CrossRef PubMed.
- C. S. Stancu, G. M. Sanda, M. Deleanu and A. V. Sima, Probiotics determine hypolipidemic and antioxidant effects in hyperlipidemic hamsters, Mol. Nutr. Food Res., 2014, 58, 559–568 CrossRef CAS PubMed.
- N. Martin Ask, M. Leung, R. Radhakrishnan and G. P. Lobo, Vitamin A Transporters in Visual Function: A Mini Review on Membrane Receptors for Dietary Vitamin A Uptake, Storage, and Transport to the Eye, Nutrients, 2021, 13, 3987 CrossRef CAS PubMed.
- M. Yaman, J. Çatak, H. Uğur, M. Gürbüz, İ. Belli, S. N. Tanyıldız, H. Yıldırım, S. Cengiz, B. B. Yavuz, C. Kişmiroğlu, B. Özgür and M. C. Yaldız, The bioaccessibility of water-soluble vitamins: A review, Trends Food Sci. Technol., 2021, 109, 552–563 CrossRef CAS.
- G. Neale, B12 binding proteins, Gut, 1990, 31, 59–63 CrossRef CAS PubMed.
- C. H. Halsted, Absorption of water-soluble vitamins, Curr. Opin. Gastroenterol., 2003, 19, 113–117 CrossRef CAS PubMed.
- I. H. Rosenberg and J. Selhub, Folate and vitamin B12 transport systems in the developing infant, J. Pediatr., 2006, 149, S62–S63 CrossRef CAS.
- N. Swiatlo, D. L. O'Connor, J. Andrews and M. F. Picciano, Relative Folate Bioavailability from Diets Containing Human, Bovine and Goat Milk, J. Nutr., 1990, 120, 172–177 CrossRef CAS PubMed.
- M. Yaman, Ö. F. Mızrak, J. Çatak and H. S. Sargın, In vitro bioaccessibility of added folic acid in commercially available baby foods formulated with milk and milk products, Food Sci. Biotechnol., 2019, 28, 1837–1844 CrossRef CAS PubMed.
- M. Verwei, K. Arkbåge, H. Mocking, R. Havenaar and J. Groten, The Binding of Folic Acid and 5-Methyltetrahydrofolate to Folate-Binding Proteins during Gastric Passage Differs in a Dynamic In Vitro Gastrointestinal Model, J. Nutr., 2004, 134, 31–37 CrossRef CAS PubMed.
- T. Inatomi and M. Honma, Ameliorating effect of probiotics in a rat model of chronic kidney disease, PLoS One, 2023, 18, e0281745 CrossRef CAS PubMed.
- M. T. Liong and N. P. Shah, Optimization of cholesterol removal, growth and fermentation patterns of Lactobacillus acidophilus ATCC 4962 in the presence of mannitol, fructo-oligosaccharide and inulin: a response surface methodology approach, J. Appl. Microbiol., 2005, 98, 1115–1126 CrossRef CAS PubMed.
- I. A. Harahap and J. Suliburska, Can probiotics decrease the risk of postmenopausal osteoporosis in women?, PharmaNutrition, 2023, 24, 100336 CrossRef CAS.
- R. A. Walhe, S. S. Diwanay, M. S. Patole, R. Z. Sayyed, H. A. Al-Shwaiman, M. M. Alkhulaifi, A. M. Elgorban, S. Danish and R. Datta, Cholesterol Reduction and Vitamin B12 Production Study on Enterococcus faecium and Lactobacillus pentosus Isolated from Yoghurt, Sustainability, 2021, 13, 5853 CrossRef CAS.
- S. Doron and S. L. Gorbach, Probiotics: their role in the treatment and prevention of disease, Expert Rev. Anti-Infect. Ther., 2006, 4, 261–275 CrossRef CAS PubMed.
- J. Slavin, Fiber and Prebiotics: Mechanisms and Health Benefits, Nutrients, 2013, 5, 1417–1435 CrossRef CAS PubMed.
- J. Ji, W. Jin, S. Liu, Z. Jiao and X. Li, Probiotics, prebiotics, and postbiotics in health and disease, MedComm, 2023, 4, e420 CrossRef PubMed.
- W. Xie, Y.-S. Zhong, X.-J. Li, Y.-K. Kang, Q.-Y. Peng and H.-Z. Ying, Postbiotics in colorectal cancer: intervention mechanisms and perspectives, Front. Microbiol., 2024, 15, 1360225 CrossRef PubMed.
- K. Y. Kareem, F. Hooi Ling, L. Teck Chwen, O. May Foong and S. Anjas Asmara, Inhibitory activity of postbiotic produced by strains of Lactobacillus plantarum using reconstituted media supplemented with inulin, Gut Pathog., 2014, 6, 23 CrossRef PubMed.
- M. E. Sanders, D. J. Merenstein, G. Reid, G. R. Gibson and R. A. Rastall, Probiotics and prebiotics in intestinal health and disease: from biology to the clinic, Nat. Rev. Gastroenterol. Hepatol., 2019, 16, 605–616 CrossRef PubMed.
- J. A. Chmiel, G. A. Stuivenberg, K. F. Al, P. P. Akouris, H. Razvi, J. P. Burton and J. Bjazevic, Vitamins as regulators of calcium-containing kidney stones—new perspectives on the role of the gut microbiome, Nat. Rev. Urol., 2023, 20, 615–637 CrossRef PubMed.
- A. Ghosal, N. S. Chatterjee, T. Chou and H. M. Said, Enterotoxigenic Escherichia coli infection and intestinal thiamin uptake: studies with intestinal epithelial Caco-2 monolayers, Am. J. Physiol.: Cell Physiol., 2013, 305, C1185–C1191 CrossRef CAS PubMed.
- M. I. Bhat, K. Sowmya, S. Kapila and R. Kapila, Potential Probiotic Lactobacillus rhamnosus (MTCC-5897) Inhibits Escherichia coli Impaired Intestinal Barrier Function by Modulating the Host Tight Junction Gene Response, Probiotics Antimicrob. Proteins, 2020, 12, 1149–1160 CrossRef CAS PubMed.
- J. Zhang, Z. He, L. Liu, H. Li, T. Wang, X. Zhu, Y. Wang, D. Zhu, Y. Ning and Y. Xu, Probiotic has prophylactic effect on spatial memory deficits by modulating gut microbiota characterized by the inhibitory growth of Escherichia coli, Front. Integr. Neurosci., 2023, 17, 1090294 CrossRef CAS PubMed.
- X. Zheng, C. Xia, M. Liu, H. Wu, J. Yan, Z. Zhang, Y. Huang, Q. Gu and P. Li, Role of folic acid in regulating gut microbiota and short-chain fatty acids based on an in vitro fermentation model, Appl. Microbiol. Biotechnol., 2024, 108, 40 CrossRef CAS PubMed.
- J. Zhang, D. Cai, M. Yang, Y. Hao, Y. Zhu, Z. Chen, T. Aziz, A. Sarwar and Z. Yang, Screening of folate-producing lactic acid bacteria and modulatory effects of folate-biofortified yogurt on gut dysbacteriosis of folate-deficient rats, Food Funct., 2020, 11, 6308–6318 RSC.
- C. Liu, X. Qi, X. Liu, Y. Sun, K. Mao, G. Shen, Y. Ma and Q. Li, Anti-inflammatory probiotics HF05 and HF06 synergistically alleviate ulcerative colitis and secondary liver injury, Food Funct., 2024, 15, 3765–3777 RSC.
- S. F. Liao and M. Nyachoti, Using probiotics to improve swine gut health and nutrient utilization, Anim. Nutr., 2017, 3, 331–343 CrossRef PubMed.
- I. Khan, Y. Bai, L. Zha, N. Ullah, H. Ullah, S. R. H. Shah, H. Sun and C. Zhang, Mechanism of the Gut Microbiota Colonization Resistance and Enteric Pathogen Infection, Front. Cell. Infect. Microbiol., 2021, 11, 716299 CrossRef CAS PubMed.
- H. J. Flint, S. H. Duncan, K. P. Scott and P. Louis, Links between diet, gut microbiota composition and gut metabolism, Proc. Nutr. Soc., 2015, 74, 13–22 CrossRef CAS PubMed.
- N. Aoudia, A. Rieu, R. Briandet, J. Deschamps, J. Chluba, G. Jego, C. Garrido and J. Guzzo, Biofilms of lactobacillus plantarum and lactobacillus fermentum: Effect on stress responses, antagonistic effects on pathogen growth and immunomodulatory properties, Food Microbiol., 2016, 53, 51–59 CrossRef CAS PubMed.
- P. Markowiak-Kopeć and K. Śliżewska, The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome, Nutrients, 2020, 12, 1107 CrossRef PubMed.
- S. W. Ragsdale and E. Pierce, Acetogenesis and the wood–ljungdahl pathway of CO2 fixation, Biochim. Biophys. Acta, Proteins Proteomics, 2008, 1784, 1873–1898 CrossRef CAS PubMed.
- F. Lutgendorff, P. A. Sandström, L. M. Trulsson, L. P. Van Minnen, G. T. Rijkers, H. M. Timmerman, H. G. Gooszen, L. M. A. Akkermans and J. D. Söderholm, Probiotics maintain mucosal barrier integrity in experimental acute pancreatitis via stabilisation of tight junctions and enhancement of mucosal glutathione, Pancreas, 2007, 35, 413 Search PubMed.
- Y. Komiyama, A. Andoh, D. Fujiwara, H. Ohmae, Y. Araki, Y. Fujiyama, K. Mitsuyama and O. Kanauchi, New prebiotics from rice bran ameliorate inflammation in murine colitis models through the modulation of intestinal homeostasis and the mucosal immune system, Scand. J. Gastroenterol., 2011, 46, 40–52 CrossRef CAS PubMed.
- S. S. Yoon and J. Sun, Probiotics, Nuclear Receptor Signaling, and Anti-Inflammatory Pathways, Gastroenterol. Res. Pract., 2011, 2011, 1–16 CrossRef PubMed.
- M. Jiang, X. Zhang, Y. Zhang, Y. Liu, R. Geng, H. Liu, Y. Sun and B. Wang, The Effects of Perioperative Probiotics on Postoperative Gastrointestinal Function in Patients with Brain Tumors: A Randomized, Placebo-Controlled Study, Nutr. Cancer, 2023, 75, 1132–1142 CrossRef PubMed.
- X. Wang, P. Zhang and X. Zhang, Probiotics Regulate Gut Microbiota: An Effective Method to Improve Immunity, Molecules, 2021, 26, 6076 CrossRef CAS PubMed.
- E. R. Mann, Y. K. Lam and H. H. Uhlig, Short-chain fatty acids: linking diet, the microbiome and immunity, Nat. Rev. Immunol., 2024, 1–19 Search PubMed.
- N. Mishra, A. Garg, S. Ashique and S. Bhatt, Potential of postbiotics for the treatment of metabolic disorders, Drug Discovery Today, 2024, 29, 103921 CrossRef CAS PubMed.
- Z. Zakrzewska, A. Zawartka, M. Schab, A. Martyniak, S. Skoczeń, P. J. Tomasik and A. Wędrychowicz, Probiotics, and Postbiotics in the Prevention and Treatment of Anemia, Microorganisms, 2022, 10, 1330 CrossRef CAS PubMed.
- Y. Zhang, R. Li, Z. Xu, H. Fan, X. Xu, S. Pan and F. Liu, Recent advances in the effects of food microstructure and matrix components on the bioaccessibility of carotenoids, Trends Food Sci. Technol., 2024, 143, 104301 CrossRef CAS.
- A. Latif, A. Shehzad, S. Niazi, A. Zahid, W. Ashraf, M. W. Iqbal, A. Rehman, T. Riaz, R. M. Aadil, I. M. Khan, F. Özogul, J. M. Rocha, T. Esatbeyoglu and S. A. Korma, Probiotics: mechanism of action, health benefits and their application in food industries, Front. Microbiol., 2023, 14, 1216674 CrossRef PubMed.
- M. Massi, Effects of probiotic bacteria on gastrointestinal motility in guinea-pig isolated tissue, World J. Gastroenterol., 2006, 12, 5987 CrossRef CAS PubMed.
- A. Fusco, V. Savio, D. Cimini, S. D'Ambrosio, A. Chiaromonte, C. Schiraldi and G. Donnarumma, In Vitro Evaluation of the Most Active Probiotic Strains Able to Improve the Intestinal Barrier Functions and to Prevent Inflammatory Diseases of the Gastrointestinal System, Biomedicines, 2023, 11, 865 CrossRef CAS PubMed.
- J. R. Grider and B. E. Piland, The peristaltic reflex induced by short-chain fatty acids is mediated by sequential release of 5-HT and neuronal CGRP but not BDNF, Am. J. Physiol.: Gastrointest. Liver Physiol., 2007, 292, G429–G437 CrossRef CAS PubMed.
- O. Kanauchi, A. Andoh and K. Mitsuyama, Effects of the Modulation of Microbiota on the Gastrointestinal Immune System and Bowel Function, J. Agric. Food Chem., 2013, 61, 9977–9983 CrossRef CAS PubMed.
- A. Kuwabara, K. Tanaka, N. Tsugawa, H. Nakase, H. Tsuji, K. Shide, M. Kamao, T. Chiba, N. Inagaki, T. Okano and S. Kido, High prevalence of vitamin K and D deficiency and decreased BMD in inflammatory bowel disease, Osteoporosis Int., 2009, 20, 935–942 CrossRef CAS PubMed.
- B. Sivakumar and V. Reddy, Absorption of labelled vitamin A in children during infection, Br. J. Nutr., 1972, 27, 299–304 CrossRef CAS PubMed.
- F. Cao, L. Jin, Y. Gao, Y. Ding, H. Wen, Z. Qian, C. Zhang, L. Hong, H. Yang, J. Zhang, Z. Tong, W. Wang, X. Chen and Z. Mao, Artificial-enzymes-armed Bifidobacterium longum probiotics for alleviating intestinal inflammation and microbiota dysbiosis, Nat. Nanotechnol., 2023, 18, 617–627 CrossRef CAS PubMed.
- A. Nakkarach, H. L. Foo, A. A.-L. Song, N. E. A. Mutalib, S. Nitisinprasert and U. Withayagiat, Anti-cancer and anti-inflammatory effects elicited by short chain fatty acids produced by Escherichia coli isolated from healthy human gut microbiota, Microb. Cell Fact., 2021, 20, 36 CrossRef CAS PubMed.
- B. Le and S. H. Yang, Efficacy of Lactobacillus plantarum in prevention of inflammatory bowel disease, Toxicol. Rep., 2018, 5, 314–317 CrossRef CAS PubMed.
- A. Rodríguez-Nogales, F. Algieri, J. Garrido-Mesa, T. Vezza, M. P. Utrilla, N. Chueca, F. Garcia, M. Olivares, M. E. Rodríguez-Cabezas and J. Gálvez, Differential intestinal anti–inflammatory effects of Lactobacillus fermentum and Lactobacillus salivarius in DSS mouse colitis: impact on microRNAs expression and microbiota composition, Mol. Nutr. Food Res., 2017, 61, 1700144 CrossRef PubMed.
- E. Reboul, Proteins involved in fat-soluble vitamin and carotenoid transport across the intestinal cells: New insights from the past decade, Prog. Lipid Res., 2023, 89, 101208 CrossRef CAS PubMed.
- H. Zafar and M. H. Saier, Comparative Genomics of the Transport Proteins of Ten Lactobacillus Strains, Genes, 2020, 11, 1234 CrossRef CAS PubMed.
- A. Biswas, K. Dev, P. K. Tyagi and A. Mandal, The effect of multi-strain probiotics as feed additives on performance, immunity, expression of nutrient transporter genes and gut morphometry in broiler chickens, Anim. Biosci., 2022, 35, 64–74 CrossRef CAS PubMed.
- M. I. Abd El-Hamid, D. Ibrahim, R. I. Hamed, H. H. Nossieur, M. H. Elbanna, H. Baz, E. M. Abd-Allah, A. S. A. El Oksh, G. A. Ibrahim, E. Khalifa, T. A. Ismail and N. F. S. Awad, Modulatory Impacts of Multi-Strain Probiotics on Rabbits’ Growth, Nutrient Transporters, Tight Junctions and Immune System to Fight against Listeria monocytogenes Infection, Animals, 2022, 12, 2082 CrossRef PubMed.
- Y. Huang and Y. Zheng, The probiotic Lactobacillus acidophilus reduces cholesterol absorption through the down-regulation of Niemann-Pick C1-like 1 in Caco-2 cells, Br. J. Nutr., 2010, 103, 473–478 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2025 |